1.7. - Auwälder
Auwälder haben eine geringe Expression im PNPG (0,03%) aufgrund der Tatsache, dass die meisten der Wasserwege sehr verengt sind, wobei die Ufervegetation auf einen schmalen Randstreifen beschränkt ist. Auf der anderen Seite hat der Homem mit der Konstruktion des Staudamms und der Einrichtung von Ackerland am Rand der Wasserwege den Bereich der Auwälder noch weiter verringert. Die Auwälder fallen in den prioritären Lebensraum 91E0 - Auenwälder mit Alnus glutinosa und Fraxinus excelsior (Alno-Padion, Alnion incanae, Salicion albae).


Im PNPG gehört die überwiegende Mehrheit dieser Wälder dem Subtyp 91E0pt2 - Auen-Birkenwälder (Carici reuterianae-Betuletum celtibericae der Allianz Osmundo-Alnion und der Klasse Salici purpureae-Populetea nigrae)) an. Die Birkenwälder werden von Birke Betula celtiberica und der Weide Salix atrocinerea dominiert und anderen typischen Arten wie Erica arborea, Carex elata subsp. reuteriana, Oenanthe crocata, Calamagrostis arundinacea, Luzula sylvatica subsp. henriquesii, Osmunda regalis und Deschampsia gallaecica begleitet. Es sollte beachtet werden, dass die höheren Lagen mit Eibe (Taxus baccata) angereichert und Ilex aquifolium angereichert sind, während in den unteren Teilen Lorbeer (Laurus nobilis) und Vogelkirsche (Prunus avium) häufiger sind (HONRADO, 2003).
 |  |
 | |
 |  |
 |   |
 | |
 |  |
 |  |
In den unteren Abschnitten der Wasserläufe mit wenig geneigten Flussbetten, wo sich Alluvialböden (Boden aus Sedimentablagerung) bilden, werden die Erlenwälder (Alnus glutinosa) dominant. Allerdings sind die Erlengalerien im PNPG wegen der zerklüfteten Geomorphologie des Geländes wenig verbreitet; denn die stark geneigten Betten der Fliessgewässer verursachen torrentielle Strömungen, die an den Ufern Erosionserscheinungen auf Kosten der Sedimentablagerung hervorrufen.
Der Rio Homem setzte auch mit dem Bau von Staudämmen Schwemmlandböden unter Wasser, die aller Voraussicht nach in den heutigen Bereichen der Stauseen existierten. Die Landwirtschaft hat die oft knappen Alluvialflächen bis an den Rand der Wasserläufe genutzt (HONRADO, 2003). Erlengalerien sind im Nationalpark nur am Lima und seinen Nebenflüssen reichlich vorhanden. Diese Erlenwälder entsprechen dem Subtyp 91E0pt1 - Ripikole Erlenwälder (ALFA, 2006). Neben der Dominanz der Erle, sind in der Regel Salix atrocinerea, Fraxinus angustifolia, Laurus nobilis, Frangula alnus und Sambucus nigra sowie zahlreiche Arten von Farnen (Narcisso cyclaminei-Alnetum glutinosae). In grösseren Höhen sind diese Arten weniger häufig (mit Ausnahme von Salix atrocinerea) und die Erle wird in der Regel von der Birke begleitet, mit Deschampsia gallaecica und Galium broterianum (Gallium-broteriani Alnetum glutinosae) (HONRADO, 2003).
  | |
|  |
|  |
 | |
|  |
In Auwäldern treten auch Wasserpflanzen, amphibische und nicht arboreale Flussvegetation auf, die sich teilweise in die Lebensräume 3130pt2 - Oligotrophe Stillgewässer mit Vegetation des Hyperico elodis-Sparganion und 3260 - Fließgewässer der basalen und montanen Stufe mit Vegetation des Ranunculion fluitantis und Callitricho-Batrachion (ALFA, 2006) einfügt. Diese Vegetation wird in einem späteren Kapitel genauer beschrieben.
1.8. – Waldkiefer-Bestände
Die überwiegende Mehrheit der Wald-Kiefer (auch Schottische Kiefer genannt) (Pinus sylvestris) im PNPG sind Plantagen oder haben ihren Ursprung als Plantagen im zwanzigsten Jahrhundert. Aber es wird von der Existenz einiger kleiner Reliktkerne im östlichen Teil der Serra do Geres wie dem Vogel-Tal und dem Tal der Ribeira Matança (SERRA & CARVALHO, 1989) ausgegangen. Mendes de Almeida (1884) hat erstmals über die Existenz von Pinus sylvestris in der Sierra Gerês berichtet. Doch schon in einer Nachricht der Pfarrei São João de Campo, die von dr. GENERAL VIGÁRIO DE BRAGA (1736) geschickt wurde, bezieht sich dieser auf die Existenz von Kiefern. Da es bis zu diesem Datum keine bekannten Hinweise auf Anpflanzungen in der Serra do Gerês gibt, sollen es natürliche wilde Kiefern sein. Auf diese Weise gelten sie als die einzigen natürlichen Kiefernwälder Portugals.
 |
 |
Doch in der Vergangenheit (etwa vor 2000 bis 2500 Jahre) war Pinus sylvestris viel häufiger in den galaicominhotischen Gebirgen wie durch Pollenaufzeichnungen (BELLOT, 1950) gezeigt wird. In der Gerês konnten sie dank des xerophytishen und subkontinentalen Charakters des Südosthanges der Serra do Gerês und der Morphologie des Biotops (Grund eingeschlossener Täler), der derzeit von der Spezies besetzt ist und ihr Zuflucht vor Feuer gewährte, überleben (HONRADO 2003). Die Pinus sylvestris Wälder ordnen sich innerhalb der Klasse Pinus-Juniperetea ein, welche die natürliche potentielle Vegetation der oromediterranen und orosubmediterranen silikatreichen iberischen Gebirge bereitstellt (Rivas-Martinez et al., 1987).
2. – Gebüsche
Die allgemeine Zerstörung der Baumvegetation begünstigte die Ausdehnung der Gebüsche und Heiden, unter denen die Ginstergebüsche aus der Klasse (Cystostea scopario-striati) und die Erika- und Stechginsterheiden aus der Klasse (Calluno-Ulicetea) überwiegen.
In der Kartographie der Natürlichen Habitate des PNPG wurde von CIBIO & FCUP (2007) folgende Einteilung der Gebüsche und Heiden vorgenommen:
1. Montane Gebüsche (über 1000 m Höhe)
1.1. Montane Gebüsche mit Felsvorsprüngen
1.2. Montane Gebüsche ohne Felsvorsprünge
2. Colline Gebüsche (unter 1000 m Höhe)
2.1. Colline Gebüsche mit Felsvorsprüngen
2.1.1. Colline Gebüsche mit Felsvorsprüngen auf syntektonischen Graniten
2.1.2. Colline Gebüsche mit Felsvorsprüngen auf Schiefer und posttektonischem Granit
2.2. Colline Gebüsche ohne Felsvorsprünge
2.2.1. Colline Gebüsche ohne Felsen auf syntektonischen Graniten
2.2.2. Colline Gebüsche ohne Felsen auf Schiefer und posttektonischen Graniten
2.1. – Montane Gebüsche mit Felsvorsprüngen
Die Montanen Gebüsche mit Felsvorsprüngen sind überall in den höchsten Teilen des Parks zu finden, aber in der Serra do Gerês dominieren sie fast vollständig. Diese Gebüsche bestehen aus Heiden (Erica australis subsp. aragonensis) (Klasse Calluno-Ulicetea) und aus Geissklee-Gemeinschaften des Cytiso striati-Genistetum polygaliphyllae, die praktisch auf die unmittelbare Nähe von Wasserläufen beschränkt sind (HONRADO, 2003).
Die Heiden werden von Erica australis subsp. aragonensis dominiert und haben die übliche Präsenz von Erica umbellata, Pterospartum tridentatum subsp. cantabricum, Pseudarrhenatherum longifolium, Ulex minor und Carex asturica (Carici asturicae-Ericetum aragonensis). Diese Heiden entsprechen dem Lebensraum 4030pt3 - Europäische trockene Heiden - Subtypus pt3 der mediterranen nicht litoralen Heidekraut-, Stechginster- und Zistrosenheiden.
In der südlichen altimontanen-Zone (über 1100 m) der Serra do Gerês, wird Erica australis subsp. aragonensis von Juniperus communis subsp alpinus, Thymelaea broteriana und Carex asturica begleitet (Gemeinschaft von Juniperus communis subsp. alpina und Erica australis subsp. aragonensis). In Portugal finden sich Heiden mit Wacholder nur in der Serra do Gerês und in der Serra da Estrela, wo sie die letzten Überreste einer Heidegemeinschaft einer ehemals breiteren eiszeitlichen Verbreitung bilden (HONRADO, 2003) Diese Heiden entsprechen dem Lebensraumtyp 4030pt4 - Geresiensische und Estrelensische Heiden-Wacholder-Gemeinschaft (ALFA, 2006).
 |  |
 |  |
 |
Ginstergebüsche sind heute, da sie einen tieferen Boden benötigen, in den Gebieten mit Felsvorsprüngen sehr selten. Die typische Zusammensetzung dieser Ginstergebüsche besteht aus: Genista florida subsp. polygaliphylla, Cytisus striatus, Erica arborea, Pteridium aquilinum und Agrostis x fouilladei (HONRADO, 2003).
 |  |
 |  |
 |
In Lichtungen von Gebüschen und in breiten Felsspalten, die der Sonneneinstrahlung und dem Wind ausgesetzt sind, treten in mittleren bis hohen Höhen xerophytische Formationen mit mehrjährigen Gräsern auf (Stipa gigantea, Avenula sulcata subsp. sulcata, Agrostis curtisii, Festuca elegans, Pseudarrhenatherum longifolium (Pseudarrhenathero longifolii-Stipetum giganteae) (HONRADO, 2003). Diese Vegetation wird als prioritärer Lebensraum eingestuft - 6220* Gräser und Annuelle der Klasse Thero-Brachypodietea, genauer des Subtypus 6220pt4 der Mehrjährigen silikolen Hochrasengesellschaften aus der Klasse: Stipo giganteae-Agrostietea castellanae (ALFA, 2006).
  |  |
  |   |
 |
In den Lichtungen der Gebüsche und in den Felsvorsprüngen finden sich noch oroiberische Rasen von Festuca indigesta (Lebensraum 6160), des Subtypus 6160pt2 - Acidophile niederliegende Gebüsche des gemäßigten und mediterranen Klimas (Hieracio castellani-Plantaginion radicatae). Diese Vegetation wird von kleinen Sträuchern (Chamaephyten) und xerophilen Gräsern, Pionieren von Skelettböden oder Felsspalten, dominiert (ALFA, 2006). HONRADO (2003) hat im PNPG vier Pflanzengemeinschaften aus der Klasse Festucetea indigestae identifiziert, die mit diesem Habitat assoziiert sind:
- Minuartio recurvae-Silenetum acutifoliae in Erdspalten von exponierten Felsvorsprüngen mit Festuca summilusitana, Sedum brevifolium, Allium ericetorum, Ornithogalum concinnum, Solidago virgaurea und Silene acutifolia.
 |  |
 |  |
  |   |
- Arenario Querioidis-Sedetum brevifoli auf Kies von konvexen Mikrotopographien mit Arenaria querioides, Sedum brevifolium, Agrostis truncatula subsp. commista und Tulipa sylvestris subsp. australis.
  | |
 |  |
- Armerio humilis-Agrostietum commistae auf sandigen Leptosolen von flachen oder konkaven Mikrotopographien und auf Skelettböden in der Nähe von Felsnasen mit Agrostis truncatula subsp. Commista, Ornithogalum concinnum, Armeria humilis subsp. humilis, Molineriella laevis und Sedum brevifolium.
| |
 |  |
|
- Plantagini radicatae-Armerietum odoratae auf Leptosolen aus syntektonischen Graniten mit Agrostis truncatula subsp. commista, Armeria humilis subsp. odorata, Plantago radicata, Ornithogalum concinnum.
| |
  | |
In felsigen Aufschlüssen finden wir felsige Vegetation des Lebensraums 8220 - Kieselige Felsnasen mit chasmophiler Vegetation, genauer gesagt im Subtyp 8220pt1 - Silikat-Gesteine mit chasmophilen Gemeinschaften (Klasse Asplenietea trichomanis) (ALFA, 2006).
Im PNPG erscheint die Assoziation Phalacrocarpo oppositifolii-Silenetum acutifoliae mit der folgenden charakteristischen Pflanzenkombination: Saxifraga spathularis, Phalacrocarpum oppositifolium, Silene acutifolia, Silene marizii und gelegentlich Sedum hirsutum, Hieracium amplexicaule und Murbeckiella boryi (HONRADO, 2003).
  |   |
|   |
 |  |
  |
In den höheren Lagen zeigen die Felsformationen einen recht seltenen, von Echinospartum ibericum beherrschten Lebensraumtyp, den Habitat 4090 - endemische oromediterrane Heiden mit dornigen Ginstern (ALFA, 2006).

Polster von Echinospartum ibericum (Serra da Estrela, Portugal)

2.2. - Montane Gebüsche und Heiden ohne Felsvorsprünge
Die Montanen Gebüsche und Heiden ohne Felsvorsprünge, die sich auf die beiden Hochebenen des PNPG konzentrieren, weisen eine geringere Expression auf als die im vorherigen Punkt beschriebenen Gemeinschaften. Es gibt nur wenige Unterschiede in den Vegetationstypen, die sich wie folgt zusammenfassen lassen:
- Eine größere Abundanz von Ginstergebüschen, da der Boden tiefgründiger ist, und das Vorkommen einer Ginstergemeinschaft im äußersten Nordosten des Parks, die durch das völlige Fehlen von Cytisus striatus und seine Substitution durch C. multiflorus und C. scoparius (Assoziation Cytiso scoparii-Genistetum polygaliphyllae) charakterisiert ist. (HONRADO, 2003).




- Anwesenheit von mesophytischen Rasen, dominiert von Arrhenatherum elatius subsp. baeticum, Agrostis x fouilladei, Armeria x francoi, Rumex acetosella subsp. angiocarpus und Hypochaeris radicata (Armerio francoi-Arrhenatheretum baetici), in den Ginsterlichtungen auf dem Planalto da Mourela und im Tourém-Tal (HONRADO, 2003). Diese Vegetation bildet einen Teil des Habitats 6220pt4 - Mehrjährige silikatliebende Hochgräser (ALFA, 2006).
  | |
 |  |
2.3. - Colline Gebüsche und Heiden mit Felsformationen auf syntektonischen Graniten
Die collinen Gebüsche mit Felsvorsprüngen auf syntektonischen Graniten haben eine geringe Expression im PNPG (2,3% Fläche). Sie werden dominiert von dem Stechginster Ulex europaeus subsp. latebracteatus des Ulicetum latebracteato-minoris (HONRADO, 2003). Die Stechginster, gekennzeichnet auch durch eine hohe Präsenz von atlantischen Elementen wie Daboecia cantabrica, Erica cinerea, Pseudarrhenatherum longifolium und Ulex minor ordnen sich bei den Stechginster- und Galizisch-Portugiesischen küstenabgewandten Heidekraut-Stechginster Heiden (4030pt2) ein (ALFA, 2006).
 | |
 |  |
| |
Es gibt auch einige von dem Geissklee Cytisus striatus dominierte Ginstergebüsche, in denen Erica arborea, Ulex minor, Pteridium aquilinum, Halimium lasianthum subsp. alyssoides und Agrostis x fouilladei (Ulici latebracteati-Cytisetum striati) ebenfalls häufig vorkommen (HONRADO, 2003).
   | |
 |  |
|   |
| |
Auf den Lichtungen von Ginstergebüschen können auf sandigem Boden und granitischen Leptosolen an wenig geneigten Hängen Thymianheiden mit Schillerndem Thymian (Thymus caespititius), Agrostis truncatula subsp. commista und Sedum brevifolium (Sedo anglici-Thymetum caespititii) angetroffen werden. (HONRADO, 2003). Diese Vegetation entspricht dem Lebensraum 8230 pt1 - Galaico-portugiesische Thymianheiden (ALFA, 2006).


In den Felsaufschlüssen existiert rupikole Vegetation, die von Silene acutifolia dominiert und in der Regel von Sedum hirsutum, Holcus gayanus und Umbilicus rupestris (Sileno acutifoliae-Holcetum gayani) begleitet wird (HONRADO, 2003). Diese Vegetation ist Teil des Lebensraums 8220pt1 - Silikatreiche Felsaufschlüsse mit chasmophytischer Vegetation (ALFA, 2006).




2.4. - Colline Gebüsche und Heiden mit Felsformationen auf Schiefer und posttektonischen Graniten
Die collinen Gebüsche mit Felsen auf Tonschiefer und posttektonischem Granit besitzen einen erhebliche Ausdehnung in den unteren Bereichen der Gebirge von Gerês und Peneda (13% der Fläche des Parks). Diese Wälder sind durch das Fehlen von Stechginster (Ulex europaeus subsp. latebracteatus) gekennzeichnet und werden von Erica umbellata, Pterospartum tridentatum subsp. cantabricum, Erica cinerea, Ulex minor und Pseudarrhenatherum longifolium (Halimio alyssoidis-Pterospartetum tridentatae) dominiert.
| |
 | |
|
In Schieferbereichen gibt es ausser Erica umbellata eine Kodominanz von Genista triacanthos mit einer üblichen Präsenz von Ulex minor, Carex pilulifera und Agrostis curtisii (Carici piluliferae-Genistetum triacanthi) (HONRADO, 2003). Diese Vegetation ist im Habitat 4030pt3 - küstenferne mediterrane Heidekraut-, Ginster- und Heidekraut-Zistrosenheiden (ALFA, 2006). In dieser Vegetation, der Assoziation Ulici latebracteati- Cytisetum striati, sind Ginster relativ selten. Sie unterscheidet sich von der im vorhergehenden Abschnitt beschriebenen lediglich durch einen größeren Anteil von Ulex europaeus subsp latebracteatus - und in den xerothermischeren Bereichen von Arbutus unedo (HONRADO, 2003).
|  
Genista triacanthos
|
|  |
| |
|  |
In den Lichtungen zwischen Heidekraut kann man zusätzlich zu dem Thymian des Sedo anglici-Thymetum caespititii auch in größerer Fülle die Gemeinschaft des Sedo pruinati-Thymetum caespititi finden. Die häufigste Florenkombination umfasst Thymus caespititiius, Sedum pruinatum, Centaurea und S. brevifolium aristata subsp. geresensis (HONRADO, 2003). Dies entspricht der Vegetation des Lebensraums 8230pt1 - Galizisch-Portugiesische Thymianheiden (ALFA, 2006).





In den Lichtungen der Gebüsche und in breiten Felsspalten an sehr dem Sonnenlicht und Wind exponierten Stellen treten gelegentlich xerophytische mittel- bis hochwüchsige Formationen dominiert durch ausdauernde Gräser (Stipa gigantea, Avenula sulcata subsp. sulcata, Agrostis curtisii, Festuca elegans, Pseudarrhenatherum longifolium, Agrostis truncatula subsp commista, Agrostis x fouilladei und Endemismus Aurea limbata subsp. geresensis (Pseudarrhenathero longifolii-Stipetum giganteae) auf (HONRADO, 2003). Diese Vegetation ist Teil des prioritären Lebensraums 6220 - Substeppen von Gräsern und Annuellen der Klasse Thero- Brachypodietea, insbesondere des Subtyps 6220pt4 - Rasen von mehrjährigen silicíkolen hohen Gräsern (ALFA, 2006).
|  |
| |
| |
|  |
In den Felsformationen existiert eine von Silene acutifolia dominierte rupikole Vegetation, die meist von Sedum hirsutum, Holcus gayanus und Umbilicus rupestris (Sileno acutifoliae-Holcetum gayani) begleitet wird (HONRADO, 2003). Diese Vegetation befindet sich im Lebensraum 8220pt1 - Silikatreiche Felsaufschlüsse mit chasmophytischer Vegetation (ALFA, 2006).
2.5. - Colline Gebüsche und Heiden ohne Felsen auf syntektonischen Graniten
Die Hügel ohne Felsvorsprünge auf syntektonischen Graniten sind auf die Gebirge Serra Amarela und Soajo und das tal des Rio Salas (Mourela) konzentriert und besetzen rund 13% der Fläche. Diese Vegetation ist identisch mit derjenigen, die für die collinen Hügel mit Felsvorsprüngen auf syntektonischen Graniten beschrieben wurde. Die Hauptunterschiede liegen in der Abwesenheit von rupikoler Vegetation und der Gegenwart von xero-mesophytischen Gräsern, die von Arrhenatherum elatius subsp. baeticum, Agrostis castellana, Agrostis x fouilladei, Armeria beirana, Centaurea limbata subsp. geresensis und Hypericum linariifolium (Armerio baetici-Arrhenatheretum beiranae) in den Lichtungen von Ginstern, auf Hügeln mit steilen Hängen, im mittleren Talabschnitt des Flusses Lima (HONRADO, 2003). Diese Vegetation passt in den Lebensraum 6220pt4 - Mehrjährige silicikole Rasen mit hohen Gräsern (Klasse: Stipo giganteae-Agrostietea castellanae) (ALFA, 2006).
Ein weiterer Unterschied, aber nur im Rio Salas-Tal, ist die größere Häufigkeit der Geissklees Cytisus multiflorus (Ulici latebracteati-Cytisetum striati) (HONRADO, 2003).
|  |
|  |
|   |
  |
2.6. - Colline Gebüsche ohne Felsen auf Schiefer und posttektonischen Graniten
Die collinen Hügel ohne Felsvorsprünge auf Schiefer und post-tektonische Granitsteinen sind im PNPG-Gebiet etwas verstreut und besetzen etwa 10% der Fläche. Diese Vegetation ist identisch mit derjenigen, die für die collinen Hügel mit felsigen Aufschlüssen auf post-tektonischen Graniten beschrieben wurde. Nur hat sie natürlich keine rupikole Vegetation und auch keine xerophytischen Rasen mit Stipa gigantea (Pseudarrhenathero longifolii - Stipetum giganteae).
3. - Agro-forstwirtschaftliches Mosaik
Das agro-forstwirtschaftliche Mosaik besteht im Wesentlichen aus bewässerten Wiesen und Heuwiesen (lameiros) sowie Streifen mit von Eichen dominierten Laubbäumen. Wiesen oder Heuwiesen werden meist mit einer dünnen Schicht Wasser ("lima") bewässert, die Frostschäden an Pflanzen (im Winter) verhindert. Aufgrund seiner floristischen Zusammensetzung können wir die regelmäßig gemähten und selten beweideten eingezäunten Wiesen (Allianz Arrhenatherion) von den gemähten und gleichzeitig beweideten Wiesen (Allianz Cynosurion) unterscheiden. Die regelmäßig gemähten und selten beweideten Weiden bilden den Lebensraum 6510 - Meso-hygrophile Heuwiesen (Alopecurus pratensis, Sanguisorba officinalis) (ALFA, 2006).

Alopecurus pratensis
| 
Sanguisorba officinalis
|
Nach HONRADO (2003) können wir innerhalb dieser unterscheiden:
- Die Gesellschaft des Agrostio fouilladei-Arrhenatheretum bulbosi, die fast im gesamten PNPG-Gebiet vorkommt, dominiert von Arrhenatherum elatius subsp. bulbosum mit Dactylis glomerata, Holcus lanatus, Anthoxanthum odoratum, Agrostis x fouilladei und Centaurea nigra subsp. rivularis.
- Eine Gemeinschaft mit Arrhenatherum elatius subsp. bulbosum und Agrostis capillaris in den niedrigeren Höhenlagen des Parks.
Die Weiden, die häufiger Mahd und Beweidung ausgesetzt sind und sich im Mourela-Plateau und im Tourém-Tal konzentrieren, werden von Cynosurus cristatus, Holcus lanatus, Anthoxanthum odoratum, Hypochoeris radicata, Trifolium repens, Centaurea rivularis und Chamaemelum nobile dominiert. Aufgrund der intensiven Nutzung besitzen sie eine geringere floristische Diversität und mehr nitrophile Pflanzen (Chamaemelum nobile, Ranunculus repens, Trifolium repens) als die Agrostio-Arrhenatheretum bulbosi-Wiesen (HONRADO, 2003). Da selbst diese Wiesen im PNPG jedoch keiner sehr intensiven Beweidung unterliegen, weisen auch sie eine hohe Biodiversität auf, und es finden sich wichtige Arten wie die Narcissus pseudonarcissus subsp. nobilis, Serapias lingua, Paradisea lusitanica und Arnica montana subsp. Atlantica.
  |   |
 |  |
 |  |
 | |
 |  |
 |   |
Wenn die Wiesen oder ein Teil von ihnen feuchter sind, werden sie von Binsen (Juncus spp.) statt der Gräser dominiert. Wenn nur eine geringe Vernässung stattgefunden hat, dominieren Juncus effusus und Deschampsia gallaecica (Deschampsio gallaecicae-Juncetum effusi). Im Falle von stärkerer Vernässung dominiert Juncus acutiflorus (Peucedano lancifolii-Juncetum acutiflori) (HONRADO, 2003). Die Wiesen mit Juncus ordnen sich in dem Lebensraum 6410 - Molinia-Wiesen auf kalkhaltigen, antorfigen und tonig-schlammigen Böden, genauer gesagt, dem Subtyp 6410pt2 - acidophile Binsenbestände mit Juncus acutiflorus, J. acutiflorus und/oder J. effusus (ALFA 2006 ), ein. In feuchten Böden in der Nähe der Wänden, Zäunen, Wasserleitungen oder verlassenen Wallungen finden sich andere Arten: Eupatorium cannabinum, Geranium robertianum, Chaerophyllum temulum und Mentha suaveolens. Diese Vegetation ist Teil des Lebensraumes 6430 - hohe hygrophile Grasgemeinschaften an basalen Rändern der montanen bis alpinen Stufe, genauer gesagt, der Subtypus 6430pt1 - Meso-hygrophile megaphorbische escionitrophile Vegetation (Galio aparines-Alliarietalia petiolatae p.p. max) (ALFA, 2006).
 |  |
 | |
  |  |
  |  |
An den Rändern der Feuchtwiesen gehören die meisten Laubbäume dem Lebensraum 9230 - Galizisch-Portugiesische Eichenwälder von Quercus robur und Quercus pyrenaica (ALFA, 2006) an. Mit den Eichen erscheinen auch die comophytischen schattenliebenden sowie epiphytischen Gemeinschaften, die dem Lebensraum 8220pt3 – Biotope comophytischer esciophiler sowie epiphytischer Pflanzengemeinschaften, (Klasse: Anomodonto-Polypodietalia) angehören (ALFA, 2006).
4. Aquatische- und Nicht-arboreale Ufervegetation
Die nicht-arboreale Vegetation der Wasserläufe des PNPG umfasst die folgenden Gemeinschaften:
- Die amphibische Pioniervegetation von Therophyten (Isoeto-Nanojuncetea) dominiert, häufig in Ufern von Wasserläufen, Stauseen und temporären Pfützen.
- Die ausdauernden amphibischen Gemeinschaften der Klasse Isoeto-Littorelletea, typisch für mehr oder weniger dauerhafte Pfützen und Stauwasser von kleinen Wasserläufen.
- Formationen von Sumpfpflanzen (Helophyten) (Klasse Phragmito-Magnocaricetea), häufig an den Rändern und in steinigen oder sandigen Betten von Strömen und in Kanälen.
- Die an Bryophyten reiche Quellvegetation der Klasse Montio-Cardaminetea, die im Gebiet eine beträchtliche phytozönotische Vielfalt aufweist.
Die therophytische amphibische Vegetation wird durch Bryum alpinum, Illecebrum verticillatum, Molineriella laevis, Juncus capitatus und Holcus gayanus dominiert. Es sind die briograminóides Gemeinschaften des Holco gayani-Bryetum alpini, die Formationen von sandigen, saisonal durchnässten Böden des Periballio laevis-Illecebretum verticillati der Frühlings- und Sommer-Phänologie, Gemeinschaften in montanen Regionen Gnaphalio uliginosi-Spergularietum capillaceae und Formationen in mesomediterrânicas Bereichen von Gemeinschaften von “Illecebrum verticillatum und Cyperus flavescens” (mit spätsommerlichen und herbstlichen Optima) identifiziert (HONRADO, 2003).
Die ausdauernden amphibischen Pflanzengemeinschaften sind im Lebensraum 3130 enthalten - Stehende, oligotrophe und mesotrophe Gewässer, mit Vegetation der Littorelletea uniflorae und/oder Isoëto-Nanojuncetea, konkreter im Subtyp 3130pt2 - Oligotrophe stehende Gewässer mit Vegetation des Hyperico elodis-Sparganion und im Lebensraum 3260 - Wasserläufe der basalen und montanen Stufe mit Vegetation des Ranunculion fluitantis und Callitricho-Batrachion (ALFA, 2006).
 |  |
 |  |
 |  
(Cyperus flavescens)
|
Im PNPG ist diese Vegetation in zwei Arten von Phytozönosen unterteilt (HONRADO, 2003):
i) Gesellschaften mit Ranunculus ololeucos oder R. omiophyllus, die die tiefsten Stellen in Kanälen, Pfützen und Staudämmen besiedeln (Fontinalo antipyreticae-Ranunculetum ololeuci und CB Ranunculus omiophyllus).


ii) von Potamogeton polygonifolius dominierte Gemeinschaften mit der üblichen Präsenz von Juncus bulbosus, Baldellia alpestris, Hypericum elodes und Ranunculus omyophyllus, die Kanäle mit langsamen und flachen Gewässern besiedeln (Hyperico elodis-Potametum oblongi).
 | |
  |  |
  |   |
(i) von Doldengewächsen wie Apium nodiflorum und Oenanthe crocata dominierte Gemeinschaften und von Gräsern wie Glyceria declinata (Glycerio declinatae-Apietum nodiflori - in Kanälen mit an Nitraten angereichertem Wasser und Glycerio declinatae-Oenanthetum crocatae - an den Ufern und Sandbänken der permanenten Gewässer).






ii) von Carex elata subsp. reuteriana, Galium broterianum und Viola palustris dominierte Gemeinschaften, die die steinigen Ränder von kleinen und mittelgroßen Wasserläufen (Gallio broteriani-Caricetum broterianae) besiedeln.



Nach (HONRADO, 2003) kann die Quell-Vegetation unterteilt werden in:
i) an fliessende, oligotrophe und gut mit Sauerstoff angereicherte Gewässer in nemoralen und peri-nemoralen Umgebungen gebundene Pflanzengemeinschaften, absolut dominiert von Chrysosplenium oppositifolium, in denen auch häufig Cardamine flexuosa, Epilobium obscurum und Stellaria alsine und mindestens zwei Dutzend Arten von Bryophyten (Cardamino flexuosae-Chrysosplenietum oppositifolii) erscheinen.
 | |
 |  |
 |
Viele Moose (Bryophyten)
|
ii) Heliophile Gemeinschaften, die flache Teiche kolonialisieren und von Ranunculus omiophyllus und Montia amporitana (Ranunculetum omiophylli) dominiert werden.

iii) Heliophile Formationen, die Kanäle mit oligotrophem flachem und langsam fliessendem Wasser kolonialisieren und von Montia amporitana und Stellaria alsine dominiert werden (CB Montia amporitana, Stellaria alsine - [Montio-Cardaminetalia]).
iv) Pflanzengemeinschaften mit rupikolen Appetenzen, die felsige Hänge mit oligotropher Wasserversorgung kolonisieren und von Saxifraga lepismigena dominiert werden; mit einer großen Fülle und Vielfalt von Moosen, wovon besonders charakteristisch sind: Scapania undulata und Marsupella emarginata (Caricion remotae: Saxifragetum lepismigenae).





v) Pflanzengemeinschaften an der Basis von Felsböschungen oder schattigen Granitwänden, mit leicht nitrifiziertem Spritzwasser, dominiert von Sibthorpia europaea, Sagina procumbens, Epilobium obscurum und Wahlenbergia hederacea (Sagino procumbentis-Sibthorpietum europaeae)



5. - Hygroturfose Komplexe
Die hygroturfosen Komplexe befinden sich in der montanen Zone des Parks und besetzen 336 ha. Diese Komplexe entsprechen in der Regel einem sehr unterschiedlichen Mosaik aus wasser- und feuchtigkeitsliebender Vegetation. Im Zentrum hygroturfoser Systeme stehen in der Regel mehr oder weniger tiefe und nichtaustrocknende Teiche mit amphibischen graminoiden von Antinoria agrostidea subsp. natans, Glyceria declinata, Juncus bulbosus, Callitriche stagnalis und Eleocharis multicaulis dominierten Gemeinschaften (Glycerio-Antinorietum agrostideae). Diese Gemeinschaften ordnen sich häufig im Mosaik mit von Ranunculus ololeucos dominierten Gemeinschaften (Fontinali antypireticae-Ranunculetum ololeuci) an.
  |   |
|  |
  |
In den Kanälen mit langsamen, flachen Gewässern von hygroturfosen Systemen gibt es noch eine Gemeinschaft dominiert von kleinen Helophyten und Hydrophyten einschließlich Potamogeton polygonifolius, Juncus bulbosus, Baldellia alpestris und Hypericum elodes (Hyperico elodis-Potametum oblongi). Diese Vegetation ist im Habitat 3130 - Stehende, oligotrophe und mesotrophe Gewässer, mit Vegetation der Littorelletea uniflorae und/oder Isoëto-Nanojuncetea, speziell im Subtyp 3130pt2 - Oligotrophe stehende Gewässer mit Vegetation Hyperico elodis-Sparganion (ALFA, 2006) enthalten.

Bereiche in unmittelbarer Nachbarschaft eines Sees oder mit permanentem Sickerwasser werden durch Pioniergemeinschaften mit Rhynchospora alba, Eleocharis multicaulis und/oder Lycopodiella inundata dominiert (Eleocharito multicaulis-Rhynchosporetum albae). Diese Vegetation erscheint im Habitat 7150 - Torfmoor-Schlenken mit Schnabelbinsen-Gesellschaften (Rhynchosporion) (ALFA, 2006). Es erscheinen zwei Vegetationstypen, der eine wird von Lycopodiella inundata, der andere von Menyanthes trifoliata dominiert.
Bei Akkumulation von organischem Material tritt die Gemeinschaft des Arnicetum atlanticae von Arnica montana subsp. atlantica, mit verschiedenen Seggen (hauptsächlich Carex panicea), Eriophorum angustifolium und Sphagnum-Moosen auf. Die Bedeckung mit Moosen der Gattung Sphagnum kann fast vollständig sein. Anscheinend werden diese Formationen in den hygroturfosen Systemen durch die Beweidung erhalten.
 | 
Seggen (hauptsächlich Carex panicea)
|
 | 
Sphagnum-Moose
|
Auf Mineralböden an den Rändern von Fliessgewässern und und kleinen Kanälen tritt die Assoziation Anagallido-Juncetum bulbosi innerhalb des Arnicetum atlanticae auf. Diese Assoziation zeigt insgesamt eine geringere Bedeckung mit Sphagnum-Arten als die Gemeinschaft des Arnicetum atlanticae, wobei Sphagnum auriculatum f. auriculatum der häufigste Taxon ist. Die häufigsten Gefäßarten sind Juncus bulbosus, Anagallis tenella und Wahlenbergia hederacea. Neben diesen Gemeinschaften gibt es auch eine Gemeinschaft, die von Pinguicula lusitanica dominiert wird. Diese Gemeinschaften fallen in den Lebensraum 7140 - Übergangs- und Schwingrasenmoore, genauer gesagt, die Subtyp 7140pt2 - Atlantische Moore (Gebirge im Nordwesten) (ALFA, 2006).

Sphagnum auriculatum f. auriculatum
| |
|   |
  | |
Die Entwicklung von Mooren aus der oben beschriebenen turfophilen Pioniervegetation führt zur Installation von turfophilen Mooren dominiert von Erica tetralix und Calluna vulgaris, installiert auf Moos-Polstern oder -Teppichen (Sphagnum sp. pl. und Aulacomnium palustre). Für den PGNP sind eine Assoziation des Erico tetralicis-Trichophoretum germanici und zwei Gemeinschaften mit der Glockenheide (“Erica tetralix mit Sphagnum capillifolium var. tenellum” und “Erica tetralix mit Sphagnum compactum") beschrieben. Diese Vegetationsmuster entsprechen dem Habitat 4010 - Nord-Atlantische Feuchtheiden mit Glocken-Heide (ALFA, 2006).


Am Rand von turfophilen Heiden in weniger durchnässten und Bereichen ohne Akkumulation von Torf installieren sich hygrophile Heidekraut-Stechginsterheiden im prioritären Lebensraum 4020 - Temperierte atlantische Feuchtheiden mit Erica ciliaris und Erica tetralix, insbesondere dem Subtyp 4020pt1 - Orophile Heidekraut-Stechginsterheiden. Diese unterscheiden sich von turfophileen Heiden durch die Anwesenheit von Ulex minor und der Abwesenheit (oder mit geringer Häufigkeit) von Torfmoosen (Sphagnum sp. pl..) (ALFA, 2006).
 |  |
|
(Sphagnum sp. pl..)
|
Nach HONRADO (2003) gibt es im PNPG vier Arten hygrophiler Gebüsche und Heiden:
- Montane, akzentuiert hygrophile Gebüsche und Heiden, an Rändern hygrotutfoser montaner Komplexe, dominiert von Erica tetralix und Ulex minor, mit der üblichen Präsenz von Calluna vulgaris, Agrostis hesperica und Carex binervis (Agrostio hespericae-Ulicetum minoris).
- Mäßig hygrophile montane Heiden und Gebüschen (“Ulex minor- und Cirsium filipendulum-Gemeinschaft“), die normalerweise mit der hygrophilen im vorherigen Absatz beschriebenen Gemeinschaft assoziiert auftreten, jedoch die weniger feuchten Böden als die Glockenheide-Formationen besetzen. Neben Ulex minor sind auch reichlich Halimium lasiantum subsp. alyssoides, Cirsium filipendulum, Agrostis x fouilladei und Galium vivianum vorhanden.
|   |
  | |
 |
- Montane, hygrophile, häufigen Bränden ausgesetzte Gebüsche und Heiden, dominiert von Calluna vulgaris mit der üblichen Präsenz von Potentilla erecta und Agrostis hesperica und einem durchgehenden Moosteppich mit Dominanz von Polytrichum (P. commune) und Sphagnum ( “Calluna und Polytrichum Gemeinschaft").
 | 
Agrostis hesperica
|

Polytrichum spec.
|
Sphagnum spec.
|
- Hygrophile nicht orophile Heiden und Gebüsche, die in einer geringeren Höhe als die anderen Typen auftreten, mit typischer floristischer Kombination von Erica ciliaris, Ulex minor, Carex binervis, Cirsium filipendulum, Potentilla erecta und noch Erica tetralix in nicht sehr niedriger Höhe (Cirsio filipenduli-Ericetum ciliaris).
Stark hygrophile Borstgrasrasen (port. cervunais) sind im Mosaik mit hygrophilen Erika-Stechginsterrheiden angeordnet, die den äußersten Vegetationsgürtel darstellen, der die kompensierten mit hygro-tropischen Systemen assoziierten Böden besiedelt. Die Borstgrasrasen sind Wiesen, die gewöhnlich beweidet werden und die durch die Zerstörung von hygrophilem Heidekraut durch Feuer und Überweidung entstehen. Die Borstgrasrasen werden vo nem Borstgras (Nardus stricta) dominiert, mit der üblichen Anwesenheit von Agrostis hesperica, Carex binervis, Festuca nigrescens subsp. microphylla, Juncus squarrosus, Potentilla erecta und Arnica montana subsp. atlantica (Agrostio hespericae-Nardetum strictae assoc. nova). Diese Rasen entsprechen dem prioritären Lebensraum 6230 - artenreiche Borstgrasrasen auf kieselsäurehaltigen Substraten in montanen Gebieten (und in submontanen Gebieten Kontinentaleuropas) (ALFA, 2006)
 | |
Agrostis hesperica
|  |
 |  |
| |
Die Fauna des PNPG (Nach ICNF; POPNPG)
Inventarisierung und Charakterisierung der Fauna
Durch die Vereinigung eines bemerkenswerten Florenreichtums mit einer für Portugal einzigartigen Physiographie gibt es in dem Peneda-Geres Nationalpark eine Reihe von natürlichen Lebensräumen, die eine vielfältige Faunengemeinschaft unterstützen. Die Wetterbedingungen, durch hohe Niederschläge Regime und Temperaturen im mittleren Temperaturbereich gekennzeichnet, ermöglichen einen großen Reichtum in der Vegetation und die Aufrechterhaltung der vielfältigen Lebensräume mit einer Vielzahl von Tierarten.
Bisher wurden 235 einheimische Wirbeltierarten verzeichnet, was einer grossen Faunendiversität entspricht. 204 der Arten sind auf nationaler und internationaler Ebene durch Konventionen (Berner Übereinkunft und Bonner Konvention), Richtlinien (Vogelschutzrichtlinie und Habitatrichtlinie) und spezifische Rechtsvorschriften geschützt; 71 erscheinen in der Liste der vom Aussterben bedrohten Arten des Portugiesischen Wirbeltier-Rotbuchs.
Von den Wirbellosen wurden vor kurzem 1196 Arten aus 9 Gruppen für die Region katalogisiert (POPNPG - ohne Quellenangabe). Davon 2 Arten von Schmetterlingen (Euphydryas aurinia und Callimorpha quadripunctata), 2 Käferarten (Lucanus cervus und Cerambyx cerdo) und eine Wegschnecke (Geomalacus maculosus), die im Rahmen der Arten FFH-Richtlinie geschützt sind (ICNF).
1. Wirbeltiere
1.1. - Säugetiere (Mammifera)
In Bezug auf die Säugetiere und die Gemeinschaft der Fleischfresser (Ordnung Carnivora), ist das Auftreten von Wolf (Canis lupus), Baummarder (Martes martes), Hermelin (Mustela erminea), Wildkatze (Felis silvestris) und Otter (Lutra Lutra), einer eng mit aquatischen Lebensräumen verbundenen Spezies, hervorzuheben.
Der Wolf (Canis lupus), eine bedrohte Spezies, hat auf dem Gebiet des PNPG seine höheren Populationsdichte auf der Iberischen Halbinsel und in den letzten Jahren ist die Population im PNPG stabil geblieben.

Während der Steinmarder (Martes foina) eine auf der Iberischen Halbinsel weitverbreitete Art ist, die in offenen Landschaften und als Kulturfolger selbst in Großstädten erscheint, ist der Baummarder (Martes martes) eine deutlich an Wälder gebundene Spezies, die im PGNP, an der südwestlichen Grenze seines euroasiatischen Verbreitungsgebietes, eines der wenigen für Portugal bekannten Vorkommen hat.

Auch das Hermelin (Mustela erminea) hat im PNPG eines seiner bekannten Vorkommen. Diese Art, die in den gemäßigten und subarktischen Zonen der Nordhalbkugel verbreitet ist, gelangt im PNPG ebenfalls an die südwestliche Grenze ihres eurasiatischen Verbreitungsgebietes. Für beide Arten ist der Erhaltungszustand im Rotbuch der Wirbeltiere Portugals als “unzureichend bekannt” angegeben.

Die Wildkatze (Felis sylvestris), die in Portugal den Schutzstatus „vulnerabel“ besitzt und hier sowohl in mediterranen als gemässigten Zonen vorkommt, tritt auch im PGNP auf.

Die emblematische Art des PGNP, das Reh (Capreolus capreolus), ist hier gut vertreten, und es gab eine deutliche Populationszunahme in den letzten Jahren, dem Trend der Populationen im Norden des Landes folgend.

Ausgestorben in Portugal seit 1892 und über 100 Jahre später im Parque Natural Baixa Limia Serra do Xurés (PNBLSX), der spanischen Fortsetzung des Nationalparks an der Grenze zu Portugal wieder eingeführt, ist der Iberiensteinbock (Capra pyrenaica) heute eine der wertvollsten Arten des PGNP. Die Population wird überwacht, und es ist in den letzten Jahren eine signifikante Erhöhung der Anzahl von Individuen in den besetzten Gebieten beobachtet worden. Trotzdem ist sie weiterhin als eine Spezies mit dem Status "kritisch gefährdet" im Rotbuch der Wirbeltiere von Portugal aufgeführt.

Unter den kleineren Säugetieren verschiedener Säugetierfamilien, auch als Kleinsäuger zusammengefasst, ist an erster Stelle der Pyrenäen-Desman (Galemys pyrenaicus) zu nennen, ein Iberischer Endemit aus der Familie de Maulwürfe (Talpidae, Desmanini), mit dem Schutzstatus “Vulnerabel”, und die Sumpfspitzmaus (Neomys anomalus) (Schutzstatus: “Unzureichende Informationen”), beide Arten mit semi-aquatischer Lebensweise. Des weiteren die Iberische Waldspitzmaus (Sorex granarius), eine endemische Art der Iberischen Halbinsel mit dem Status "Unzureichende Informationen".
Es muss hier noch dazu gesagt werden, dass die Einschätzungen zum Schutzstatus im Rotbuch der Wirbeltiere Portugals naturgemäß subjektiv sind, aber auch, weil systematische demographische Studien (z.B. bei den Kleinsäugern mit Hilfe von Eulengewöllen) in vielen Fällen (vor allem bei den kleineren Wirbeltieren) fehlen. Deshalb erscheint hier häufig der Schutzstatus “Unzureichende Informationen”. Außerdem ändern sich die Umweltbedingungen auf der Iberischen Halbinsel z.Zt. dramatisch, sowohl durch Waldbrände und den damit verbundenen Insektenschwund (was vor allem die Fledermäuse, aber auch andere insektivore Kleinsäuger betrifft), als auch durch den rasant fortschreitenden Klimawandel. Die Einschätzungen sind daher wahrscheinlich noch zu optimistisch und es fehlen mit Sicherheit Arten, deren Schutzstatus bereits zumindest als gefährdet angenommen werden muss.

Galemys pyrenaicus
| 
Neomys anomalus
|

Sorex granarius
| 
Gartenschläfer
|
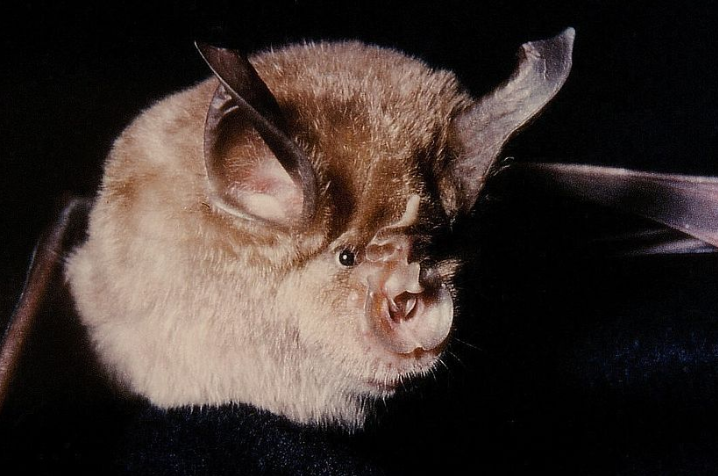
Die Fledermausgemeinschaft im PGNP besteht hauptsächlich aus Arten mit atlantischer Verbreitung, obwohl auch submediterranen Arten wie Rhinolophus euryale vorhanden sind. Es sind 15 Fledermausarten für den PNPG bekannt, davon 11 als bedroht eingeschätzt. Bemerkenswert ist der Nachweis der Mittelmeer-Hufeisennase (Rhinolophus euryale), die als "kritisch gefährdet" angesehen wird; und der Grossen Hufeisennase (Rhinolophus ferrumequinum), der Kleinen Hufeisennase (Rhinolophus hipposideros), deren Satus als “verletzbar” eingeschätzt wird. Das Große Mausohr (Myotis myotis) existiert im PNPG; aufgelistet werden auch Mopsfledermaus (Barbastella barbastellus) und Kleiner Abendsegler (Nyctalus leisleri), gut an Laubwald angepasste Arten und charakteristisch für gut entwickelte Laubhölzer, was die Bedeutung der Erhaltung der bereits selten Fleck von reifem natürlichem Wald in Portugal unterstreicht.

Mittelmeer-Hufeisennase
| 
Grosse Hufeisennase
|

Grosses Mausohr
| 
Mopsfledermaus
|
 
Kleiner Abendsegler
| |
1.2. - Vögel (Aves)
In Bezug auf die Vogelfauna sind 173 Stand- Zug- und Überwinterungsarten sowie Einzelbeobachtungen für den PNPG verzeichnet, wobei auf die ornithologische Bedeutung der Hochebene von Mourela hinzuweisen ist. Die Vielfalt der im Gebiet von Mourela vorkommenden Arten schwankt beträchtlich während des ganzen Jahres und von Lebensraum zu Lebensraum, wobei 36 der hier aufgelisteten Arten im Portugiesischen Rotbuch der Wirbeltiere erscheinen. Von dieser Gruppe gehören die folgenden zu den in ganz Portugal seltenen Arten: Circus cyaneus, Pernis apivorus Gallinago gallinago, Lanius collurio, Emberiza citrinella, Sylvia borin und Saxicola rubetra. Für die Art Saxicola rubetra ist Mourela der einzige in Portugal bekannte Nistplatz, während für Lanius collurio und Emberiza citrinella auch auf dem Plateau von Castro Laboreiro Plateau Nistplätze bekannt sind. Ebenfalls sehenswert eine besondere Erwähnung für das gesamte Gebiet des PGNP verdienen die Art Turdus philomelos , die in Portugal zum ersten Mal im Jahr 1991 als Brutvogel erschienen ist, sowie die Arten Pyrrhula pyrrhula, Anthus trivialis, Pyrrhocorax pyrrhocorax und Falco peregrinus.
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
1.3. - Kriechtiere (Reptilien)
Von den 28 in Portugal vorkommenden Reptilienarten gibt es 19 Arten im PNPG. Am bemerkenswertesten ist nach ihrem Schutzstatus das Vorkommen der Europäischen Sumpfschildkröte (Emys orbicularis), der Nordiberischen Kreuzotter (Vipera seoanei), beide mit dem Status "gefährdet", und der Schlingnatter (Coronella austriaca) und Stülpnasenotter (Vipera latastei), mit dem Status "verwundbar". Zu erwähnen sind auch die Lacerta-de-água (Lacerta schreiberi) und die Nordiberischen Kreuzotter (Vipera seoanei), ein Endemismus des Nordens der Iberischen Halbinsel, sehr selten in unserem Land, dessen Verbreitung in Portugal auf Gebiete der PNPG beschränkt ist. (Castro Laboreiro, Soajo und Tourém) und andere Populationen in der Nähe von Paredes de Coura und Montalegre und Serra do Larouco.
  | |
  | |
  | |
  | |
  |
1.4. - Lurche (Amphibien)
Von den 17 Amphibienarten, die es in Portugal gibt, gibt es im PNPG 13 Arten. Von diesen besitzen der Goldstreifen-Salamander (Chioglossa lusitanica) und der Fadenmolch (Triturus helveticus) den Schutzstatus "verletzbar" und der Iberische Scheibenzüngler (Discoglossus galganoi) den Status "fast bedroht".
 |  |
 | |
1.5. – Fische (Pisces)
Gebirgswasserläufe, oft auch Forellenbäche genannt, sind wegen ihrer für Forellen (Salmo trutta) geeigneten Eigenschaften Ökosysteme von großer Bedeutung. In der PNPG gibt es 6 Arten von einheimischen Fischen, und mindestens 5 Arten von eingeführten Fischen können, vor allem in den Stauseen angetroffen werden. Panjorca (Chondrostoma arcasii) und Europäischer Aal (Anguilla anguilla) können für ihren Erhaltungszustand "IN GEFAHR" genannt werden.
 | |
 |  |
2. – Wirbellose (Invertebraten)
Von den Wirbellosen wurden vor kurzem 1196 Arten aus 9 Gruppen für die Region katalogisiert (POPNPG - ohne Quellenangabe). Davon 2 Arten von Schmetterlingen (Euphydryas aurinia und Callimorpha quadripunctata), 2 Käferarten (Lucanus cervus und Cerambyx cerdo) und eine Wegschnecke (Geomalacus maculosus), die im Rahmen der Arten FFH-Richtlinie geschützt sind (ICNF).
  | |
 | |
 |  |
 | |
F14 Rusco aculeati-Quercetum roboris Braun-Blanquet, P. Silva et Rozeira 1956; Blechno spicant-Quercetum roboris Tüxen et Oberdorfer 1958.
|

F23 Myrtillo-Quercetum roboris Braun-Blanquet, P. Silva, Rozeira et Fontes in P. Silva, Rozeira et Fontes 1950 (holotype!).
|

F25 Linario triornithophorae-Quercetum petraeae (Rivas-Martínez, Izco et Costa ex F. Navarro 1974) F. Prieto et Vázquez 1987; Luzulo henriquesii-Quercetum pyrenaicae (F. Prieto et Vázquez 1987) F. Prieto et Vázquez 1994.
|
Auszug aus der “Karte der Natürlichen Potentiellen Vegetation Europas” (BOHN et. al. 2002)
Mit der biogeographischen, floristischen und faunistischen Beschreibung des Nationalparks Peneda-Gerês sind wir nun an das Ende des portugiesischen Abschnittes unserer botanisch-zoologischen Rundreise auf der Iberischen Halbinsel angelangt und wir verlassen das kontinentale Portugal, um unsere Rundreise in Galizien in Nordwestspanien fortzusetzen.
Die Vegetation Nordwestportugals ist jedoch so eng mit der Vegetation Galiziens verbunden, dass wir beim Eintritt nach Spanien und Galizien, ausser in der Sprache, floristisch und faunistisch zunächst kaum den Übergang von dem einem in das andere Land spüren. Was uns jedoch fehlen wird, ist die Datenbank Flora-On, die nur für Portugal besteht. Stattdessen werden wir auf andere Hilfen im Internet wie die von der spanischen Community erstellte Datenbank Anthos zurückgreifen. Und auch neu erschienene Fachliteratur wie die beiden Bände “The Vegetation of the Iberian Peninsula” (LOIDI et al., 2017) werden uns - ausser dem schon klassischen “Polunin & Smythies” (1973), zur weiteren Erforschung der ‘plant-hunting regions’ und faunistischen ‘hotspots’ auf der Iberischen Halbinsel zur Verfügung stehen.
Dennoch verspüren wir beim Eintritt in das für uns weitgehend noch unbekannte Spanien ein enthusiastisches Gefühl der Freiheit:. ein großes Land und eine neue Sprache! Sicherlich ist es eine subjektive Empfindung, aber verspüren wir das Gefühl der Freiheit nicht immer auf Reisen, wenn wir in ein neues, unbekanntes Land eintreten?
Fortsetzung des Artikels:
http://polunin01.blogspot.pt/2017/11/255c-reisestationen-serra-do-geres.html
(Fussnoten und Bibliographie: Da sich Fussnoten einschl. Bibliographie nicht ohne weiteres aus Google-Docs heraus nach Blogger übertragen lassen, können diese im veröffentlichten Originalartikel eingesehen werden -> 2.5.5 - Reisestationen (Serra do Gerês - Botanisch-Zoologische Reisen - Iberische Halbinsel)
Dennoch verspüren wir beim Eintritt in das für uns weitgehend noch unbekannte Spanien ein enthusiastisches Gefühl der Freiheit:. ein großes Land und eine neue Sprache! Sicherlich ist es eine subjektive Empfindung, aber verspüren wir das Gefühl der Freiheit nicht immer auf Reisen, wenn wir in ein neues, unbekanntes Land eintreten?
Fortsetzung des Artikels:
http://polunin01.blogspot.pt/2017/11/255c-reisestationen-serra-do-geres.html
(Fussnoten und Bibliographie: Da sich Fussnoten einschl. Bibliographie nicht ohne weiteres aus Google-Docs heraus nach Blogger übertragen lassen, können diese im veröffentlichten Originalartikel eingesehen werden -> 2.5.5 - Reisestationen (Serra do Gerês - Botanisch-Zoologische Reisen - Iberische Halbinsel)
No comments:
Post a Comment