Annexe
Annex 1 - Zur Ökologie von Nieder- und Hochmooren (Auszüge aus: WILMANNS, O. 1988)
…
Grundbegriffe
Der schwedische Geobotaniker DU RIETZ definierte (1954) ein Moor als „jede in der Natur abgegrenzte Einheit von (wenigstens großenteils) torfbildender Vegetation auf (wenigsten zeitweise) nassen Torfböden und mit einer Reihe für diese charakteristischen Arten, zusammen mit dem von dieser Vegetation seit dem Beginn der Torfbildung abgelagerten Torfe". (Obacht ist beim Studium nicht-botanischer Schriften geboten, da „Moor" zuweilen bloß „Torflagerstätte" oder auch „Boden mit > 75 % organischer Substanz" bedeutet). Zur Anhäufung unzersetzter Vegetationsreste, eben von Torf, kommt es, weil unter zeitweiliger Wasserbedeckung infolge Sauerstoffmangels weniger Humus zersetzt als nachgebildet wird.
Je mächtiger die Torfdecke, desto eher ist eine gewisse Unabhängigkeit vom geologischen Untergrund gegeben. Danach hat man die beiden Hauptmoortypen zu unterscheiden: Nieder- und Hochmoore. Während bei dünner Torfdecke das sie durchtränkende Wasser noch mit dem ionenliefernden Mineralboden in Kontakt tritt, wird dieser Stoffaustausch bei großer Torfmächtigkeit schließlich unterbrochen, es sei denn, es erfolge ein seitlicher Zustrom von mineralbodenbeeinflußtem Wasser. Ist dies nicht der Fall, so muß schließlich die Vegetation ihren Wasser- und lonenhaushalt allein aus Niederschlägen und Flugstaub, der u. a. auch Pollenkörner enthält, bestreiten; sie ist nicht mehr minerotraphent, sondern ombrotraphent und damit auch extrem oligotraphent. Ehe Ombrotrophie zustandekommt, muß also ein Stadium der Minerotrophie durchlaufen worden sein.
Hochmoore im strengen Sinne sind solche Moore oder Moorteile, die rein ombrotroph sind; sie können auch Regenwassermoore (engl. bog, schwed. mosse) genannt werden; aber auch Moore, die aus einem ombrotrophen Kern mit einem minerotrophen Rand aufgebaut sind, bezeichnet man als Hochmoore. Niedermoore (engl. fen, schwed. kärr) sind minerotrophe Moorteile
...
24. Klasse: Scheuchzerio-Caricetea nigrae, Niedermoor- und Schlenkengesellschaften
Standortsökologie und Gefährdung
Sümpfe und Niedermoore ( = Flachmoore = Kleinseggenmoore) zeichnen sich dadurch aus, daß Grund-, Quell- oder Sickerwasser den Boden langfristig durchtränkt; sie trocknen nur ganz oberflächlich ab. In Mooren kommt es dabei zu einer Hemmung der Streuzersetzung, in Sümpfen nicht; Moore bilden daher Torf als semiterrestrische Humusform, d. h. die Struktur der Bestandteile ist mikroskopisch noch erkennbar; bei Sumpfhumus ist dies nicht der Fall. Auf Torfbildung und -wachstum sei erst bei den Hochmooren näher eingegangen (s. S. 256), wo diese Prozesse den Standort stärker bestimmen als bei den Niedermooren. Diese stehen den Sümpfen noch so nahe, daß manche Gesellschaften an beiden Standorten vorkommen; wollte man das in den deutschen Namen zum Ausdruck bringen, so würden diese zu schwerfällig. Im Alpenvorland lassen sich aufgrund von Differentialarten (Sesleria varia, Gentiana verna, Pinguicula alpina u. a.) sogar innerhalb der Kalkflachmoore diejenigen Bestände, deren Wurzelboden mineralreich ist, von denen auf reinem Torf unterscheiden.
Die beiden Niedermoor-Ordnungen leben in Quellmulden, an durchsickerten Hängen, an Seen oberhalb des Großseggengürtels, auch am Rande von Hochmooren, wo sie dann mit Oxycocco-Sphagnetea-Gesellschaften in Kontakt treten und entsprechende Gradienten bilden. Die Schlenkengesellschaften, klarer: Scheuchzerietalia, sind von ihnen ökologisch abgesetzt durch stehendes, nicht sickerndes Wasser; dies führt in flachen Schlenken u. a. zu Temperaturextremen; solche kommen auch in oligotrophen Hochmoorgewässern vor.
Insgesamt sind die Nährstoffverhältnisse der Scheuchzerio-Caricetea-Gesellschaften trotz gelegentlicher leichter Überschneidungen in einzelnen Faktoren deutlich günstiger als die der Oxycocco-Sphagnetea. Manche dieser Standorte sind sicher sekundär; sie wären ohne Mahd oder Beweidung von Gesträuch oder Wald bewachsen oder doch so stark beschattet, daß die lichtliebenden Moorarten nicht gediehen.
Generell sind alle diese Standorte zivilisationsgefährdet, zumal sie kleinflächig sind, oft inmitten von landwirtschaftlich genutztem Gelände liegen und leicht von dort durch Düngung beeinflußt werden, wie man an eindringenden Naßwiesenarten erkennt; so sind sie auch meist leicht dränierbar und somit „meliorierbar".
Bei geringen Eingriffen dringen differenzierende Arten aus verwandten Gesellschaften ein, falls solche aus der Nachbarschaft einwandern können oder von Tier oder Mensch eingeschleppt werden, bei starken Veränderungen verschwinden die Moorgesellschaften völlig. Da die meisten ihrer Arten – gerade die allgemein bedrohten – keine dauerhafte Samenbank bilden (PFADENHAUER & MAAS 1987), ist eine spontane Regeneration nicht möglich. (Neben der hier angedeuteten standörtlich bedingten Variabilität kommen auch arealgeographische, sukzessionsbedingte und vegetationsgeschichtliche Differenzierungen vor.)
…
37. Klasse: Oxycocco-Sphagnetea, Hochmoorbultgesellschaften und Heidemoore
Bau und Lebensweise der Sphagnen und ihre Bedeutung für das Hochmoor
Um Entstehung und Haushalt der Hochmoore zu verstehen, muß man sich gründlich mit den Massen-Torfbildnern und damit Schlüsselarten vertraut machen, den Torf- oder Bleichmoosen, der Gattung Sphagnum. Ein Bestand von Sphagnum-Pflanzen stellt ein schwammartiges Kapillarsystem dar (s. Lehrbücher der Systematischen Botanik), das nach einer Trockenperiode von Luft, nach Regen aber von Wasser erfüllt ist. Die „Moosblättchen" bestehen zum überwiegenden Teil aus abgestorbenen, oft spiralig versteiften und sogar mit groben Poren versehenen Zellen, Hyalozyten, mit Wasserspeicherfunktion, zwischen denen, volumenmäßig zurücktretend, assimilierende Chlorozyten liegen. Die „Stämmchen" sind von einem Zylinder toter Speicherzellen umkleidet. Kapillarräume entstehen auch zwischen den meist hohlen „Blättchen" und ihrem „Ast" sowie zwischen den polstrig-gedrängt wachsenden ganzen Pflanzen. Die daraus resultierende Wasserspeicherkapazität ist artspezifisch und beträgt für S. magellanicum (= S. medium) das 21-25-fache des Trockengewichts, für S. recurvum das 14-18-fache (rund 20 bzw. 10 g/100 ccm, s. OVERBECK 1975).
Eine Regulationsmöglichkeit der Wasserabgabe besteht bei diesen Thallophyten natürlich nicht. Da die Evaporation in Hochmooren an warmen Strahlungstagen etwa der in Trockenrasen entspricht (FARBAS 1931), ist es verständlich, daß Hochmoore an relativ niederschlagsreiche und evaporationsschwache Gebiete gebunden sind (s. u.).
– Die „Schwammstruktur" einerseits und die Dichte infolge stärkerer Zersetzung andererseits bewirken, daß Sphagnum-Rasen und aus ihnen entstandener Torf wenig wasserdurchlässig und damit wenig durchsickerbar sind; der mooreigene Grundwasserspiegel kann daher kleinflächig wechseln.
Die anhaltende Durchtränkung bewirkt Luftabschluß der tieferen Schichten und entsprechende Sauerstoffarmut. Unter anaeroben Verhältnissen wird zunächst die ohnehin nur geringe Eiweißmenge abgebaut; Gasblasen aus Methan, die beim Tritt in Schlenken aufsteigen, und der Geruch nach H2S machen dies sinnfällig. Tiefwurzler können hier nicht existieren. Die Abnahme an Sauerstoff und die relative Anreicherung an Kohlenstoff und Wasserstoff stellen das erste Stadium der Inkohlung dar; Torflager, wenn auch nicht aus Sphagnen, sondern überwiegend aus Kormophyten aufgebaut, waren Voraussetzung für die Bildung von Braun- und später Steinkohle unter Druck über Jahrmillionen hin. Im Torf erhalten sich nicht nur bekanntlich schwer zersetzliche Polymere wie Lignin (und damit Holz) und Sporopollenin (und damit das äußere Sporoderm, Exospor, Exine) von Farnsporen und Pollenkörnern, sondern in geringerem Maße auch Zellulose. Die Zellwände der Sphagnen sind übrigens schwerer zersetzbar, als es reinen Zellulose-Pektin-Wänden entsprechen würde. So lassen sich die Sphagnum-Arten, die den jeweiligen Torf gebildet haben, bei geringeren Zersetzungsgraden nach dem Zeltnetz identifizieren. Moore sind daher die besten „Archive" der nacheiszeitlichen Vegetationsgeschichte.
Daß die Bleichmoose gerade für den extrem nährstoffarmen Standort Hochmoor so geeignet sind, beruht ferner auf einer weiteren Eigenart im Chemismus ihrer Zellwände: Infolge eines hohen Anteils an Polyuronsäuren wirken die Wände als Kationenaustauscher. Man wird sie als „Nährstoff-Fangorgan" und damit als ökologische Anpassung werten dürfen. Kationen, die mit Staub und Niederschlägen auf das Moor gelangen, werden rasch gegen Protonen eingetauscht, welche ihrerseits das Substrat ansäuern; dabei stellt sich ein physikalisch bestimmtes Gleichgewicht zwischen Lösung und Zellwand ein. Die Pflanze kann diese locker gebundenen basischen Kationen allmählich gegen eigene Protonen eintauschen und aktiv, entgegen der Konzentrationsgefälle, aufnehmen. Dabei wird der „Austauscher regeneriert". Der Kationengehalt des von Moosen abgepreßten Wassers kann daher stark schwanken und vom „Standortswasser" abweichen. Die Azidität des Hochmoores bleibt hoch; pH-Werte von 3-4 sind normal.
Das Wachstum der Sphagnen ist dank dieser Anpassungen erstaunlich groß: Der Längenzuwachs ist artspezifisch verschieden und kann von wenigen cm (bei den bultbildenden Sphagnen wie S. medium) bis zu etwa 50 cm/Jahr in Schlenken reichen (s. dazu DIERSSEN & DIERSSEN 1984). Solche locker und gut durchlichtet wachsenden Schlenkenpflanzen (z. B. S. recurvum) sinken auf die Seite und schieben sich so über seitliche Konkurrenten. Das Wachstum einer Hochmooroberfläche kann man auf jährlich 2-10 cm schätzen. Da die toten Pflanzenteile später zusammengepreßt werden, sind freilich für die Bildung von 1 cm Torf größenordnungsmäßig 10-20 Jahre anzusetzen (z. B. CASPARIE 1969, mit Radiokarbondatierung). Diese positive Stoffbilanz wird nach Entwässerung infolge von Torfzersetzung bei Durchlüftung negativ.
Annex 2 - Phytosoziologische Tabellen
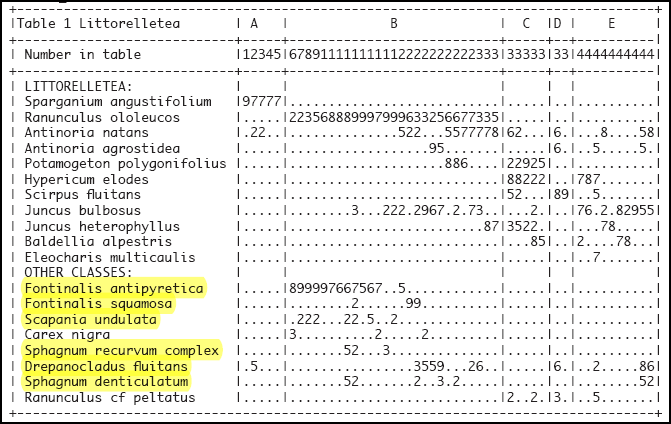
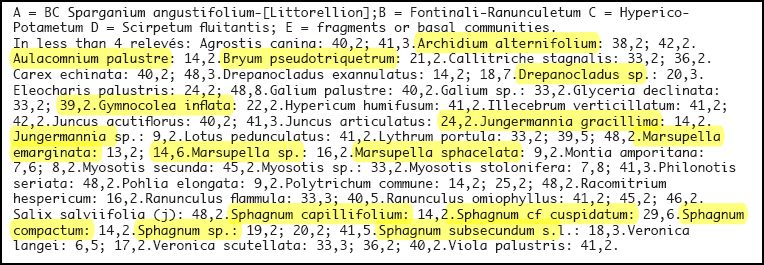
Eine synoptische Tabelle von Pflanzenaufnahmen der Vegetation der Klasse Littorelletea der Serra da Estrela findet sich bei Jan JANSEN (2011): Managing Natura 2000 in a changing world: - Radboud Universiteit (Kapitel 6, Table 1 … in der Tabelle gelb unterlegt die Flechten- und Moosarten).
Tabelle 1
Chapter 6, Table 1 Littorelletea in the Serra da Estrela
   |
Eine synoptische Tabelle von Pflanzenaufnahmen der Vegetation der Klasse Isoëto-Nanojuncetea der Serra da Estrela findet sich ebenfalls bei Jan JANSEN (2011): Managing Natura 2000 in a changing world: - Radboud Universiteit (Kapitel 6, Table 2 … in der Tabelle gelb unterlegt die Flechten- und Moosarten).
Tabelle 2
Chapter 6, Table 2 Isoëto-Nanojuncetea in the Serra da Estrela
    |
Annex 3 - Phytosoziologische Syntaxa und Habitatkorrespondenzen oder katenale Habitate in Natura 2000
Die phytosoziologischen Syntaxa der Vegetation der Serra da Estrela sind von Jan JANSEN (2002) in seinem ‘Geobotanical guide of the Serra da Estrela’ aufgelistet. Hier ein Auszug der Liste der Syntaxa zu den aquatischen Ökosystemen sowie deren Beschreibungen. Letztere sind aus den Arbeiten von RIVAS-MARTINEZ et. al. (2002): und COSTA et al. (2012) und WILMANNS, O. (1988) zusammengestellt worden.
FLOATING OR ROOTED SUBMERGED AQUATIC VEGETATION
|
Ia. FRESH-WATER VEGETATION
|

Klasse LEMNETEA: Bryo-kormophytische Süßwasser-Schwimmpflanzengemeinschaften (Lemnetalia) [kleine bis mittlere Akropleustophyten (Lemnion minoris), Mesopleustophyten von Lemniden und Riccieliden (Lemnion trisulcae) oder Hydrochariden (Lemno-Hydrocharition morsus-ranae)]. Kosmopolitisch.
Beschreibung:
Lichtgrüne Decken von Lemnaceen gehören als „Entengrütze" zum vertrauten Bild ruhiger Gewässer (Seen, Teiche, Tümpel, Gräben, Altwässer) unserer Breiten. Recht selten sind auch andere Arten mit ihnen vergesellschaftet. Die Struktur der Bestände ist einförmig: frei an oder unter der Oberfläche schwimmende Kormen, seltener Thalli bilden eine dünne einfache oder eine doppelte Schicht. Sie sind voneinander ziemlich unabhängig, erst bei dichtem Bewuchs konkurrieren sie um Raum und damit Licht.
Von Wind und Strömungen werden sie leicht verdriftet und im Strömungsschatten, z. B. zwischen Röhricht, angetrieben. Auch durch Vögel leicht verschleppbar, können sie sich rasch in Lücken innerhalb anderer Wasser- und Sumpfgesellschaften ansiedeln. Dieses „Vagabundieren" war auch Ursache dafür, daß die Klasse erst relativ spät (1955) als eigenständig erkannt worden ist. In syntaxonomisch enger und scharfer Fassung wurde sie durch TÜXEN (1974a) zunächst gründlich für Nordwestdeutschland, dann von SCHWABE-BRAUN & TÜXEN (1981) weltweit und – für die Pflanzensoziologie ungewöhnlich – auch evolutionsbiologisch behandelt.
Oft sind kleine Gruppen von Individuen oder Gesellschaftsfragmente anderer Gesellschaften, z. B. von Röhrichten, eingestreut; solche Mosaike werden als Überlagerungen aufgefaßt, die im genannten Prodromus (1981, S. 7) folgendermaßen definiert werden: „Ephemere Verzahnungen von Gesellschaften (Gesellschaftsgruppen) mit unterschiedlichem Rang in der soziologischen Progression, die keine fest zu umreißende raum-zeitliche Beziehung zueinander haben und keinen standörtlichen Gradienten aufweisen." Von Durchdringungen spricht man bei entsprechenden langfristigen Mischungen. Beide Typen von Mustern kommen als entsprechende Komplexe in der Zivilisationslandschaft mit ihren vielen Störstandorten häufig vor; die Benutzung dieser Begriffe kann oft Klarheit schaffen.
In soziologischen Aufnahmen, besonders älteren, sind häufig solche Komplexe wiedergegeben. Infolge der Artenarmut und der Kleinheit der Individuen genügen schon wenige Quadratzentimeter zur Aufnahme, bei größeren Flächen besteht die Gefahr, „verwaschene Typen" zu bekommen.
Die Lemnetea bilden Pioniergesellschaften, die über Jahrzehnte hin erhalten bleiben können (Dauer-Initialgesellschaften); ja, dichte Decken verhindern sogar das Aufkommen von Folgegesellschaften.
Die Wasserlinsendecken erfüllen also modellartig die Kriterien soziologisch einfacher Gesellschaften (s. S. 23) und stehen zu Recht am Anfang des Systems. Soziologische Primitivität bedeutet freilich nicht phylogenetische Primitivität der aufbauenden Arten, hier sogar das Gegenteil: Der Lebensraum ist für Kormophyten extrem in dem Sinne, daß er adaptive Abweichungen vom morphologischen Typus verlangt.
Charakteristische Arten: Lemna minor
|

Klasse POTAMETEA: Aquatische Frischwasser-Makrophytengemeinschaften. Verwurzelte Hydrophyten (Potametalia) [Süßwasser-Elodeide (Potamion), Brackwasser-Elodeide (Zannichellion pedicellatae), Batrachiden von fließendem Wasser (Ranunculion fluitantis), Batrachiden von stehenden Wasser (Ranunculion aquatilis) und Nymphaeiden (Nymphaeion albae)]; Nicht verwurzelte nährstoffreiche Hydrophyten (Utricularietalia) [schwimmende Utriculide (Utricularion) und Ceratophyllide (Ceratophyllion demersi)]. Kosmopolitisch.
Beschreibung:
(Die sprachlich korrekten, allerdings schwerfälligen Namen Potamogetonetea etc. sollen laut Nomenklatur-Code auf Potametea etc. verkürzt werden).
Diese Gesellschaften besiedeln nicht nur ursprüngliche, sondern in hohem Maße auch menschlich beeinflußte Gewässer und zwar sowohl lotische als auch lenitische Biotope. So ist diese Klasse häufig vertreten und dem Aspekt nach jedermann vertraut, gehören doch Seerosendecken und Laichkrautwiesen in Stillgewässern sowie Fluthahnenfußschwaden in Fließgewässern hierher. In Seen pflegen die Gesellschaften einen wasserseitigen Gürtel vor dem Röhricht aufzubauen. Dieser kann nochmals in sich gegliedert sein: Vom Ufer her bis zu etwa 4 m Wassertiefe kann die Teichrose ihre Schwimmblätter noch an die Wasseroberfläche emporschieben; dann ist nur noch submerse oder pleustische Lebensweise möglich. Unterwasserpflanzen kommen jedoch auch im ufernahen Gürtel vor, zumal wenn dieser geschädigt ist; so ist mit übergreifenden Arten zu rechnen.
Wie schwierig diese Klasse zu gliedern ist, zeigt die Vielzahl der Vorschläge; indessen lassen sie alle doch auch, wie gewohnt, Blöcke von verwandten Gesellschaften erkennen, die eben verschieden angeordnet sein können. Einige Ursachen für die praktischen und theoretischen Schwierigkeiten (s. auch WIEGLEB 1981) sind folgende: Die Wuchsorte guter Bestände sind oft schlecht zugänglich; feinstandörtliche Unterschiede und Grenzen sind häufig schwer zu erkennen; die Identifikation der oft sterilen und dazu phänotypisch ungemein variablen Arten ist oft heikel. Da die Studien vielfach nicht typologische, sondern floristische Ziele verfolgen, ist das Aufnahmematerial nicht so reich, wie es scheint. Die Interpretation von artenarmen Beständen als Sukzessionsphasen, als Relikte, als Fragmente infolge von Unterhaltsmaßnahmen und andres, aber auch die Bewertung von Wuchsformen und Vitalitätsänderungen setzt viel Erfahrung voraus. Eine stärkere Berücksichtigung des Feinmosaiks, besonders der physikalischen Faktoren Strömung, Wassertiefe und Sedimentation, zeigt daß der vermeintliche Zufall doch vielfach als Standortsabhängigkeit zu verstehen ist (Beispiel: REMY 1994).
Charakteristische Arten: Callitriche stagnalis, Potamogeton natans, Ranunculus peltatus, Ranunculus pseudofluitans.
|
AMPHIBIOUS VEGETATION OF FRESH-WATERS, SPRINGS AND FENS
|
IIa. PIONEER EPHEMERAL VEGETATION
|

Klasse BIDENTETEA TRIPARTITAE: Annuelle Pionierpflanzen an periodisch überschwemmten nährstoffreichen Stillgewässern, Seeufern und nitrophilen Flussufer-Gemeinschaften (Bidentetalia tripartitae) [feucht hoch gestört (Bidention tripartitae), austrocknenden Ufer-Tonschlämmen (Chenopodion rubri)]. Holarktisch
Beschreibung:
Das Gegenstück zu den marinen halophytischen Cakiletea bilden im Binnenland die Bidentetea. Sie kommen, wie der Monographie TÜXENS (1974a/78) zu entnehmen ist, in Europa und Ostasien vor. Standorte für die eutraphenten, Meterhöhe erreichenden und mastigen Sommerannuellen bieten sich an den Ufern von Flüssen, Gräben, nichtverdolten Dorfbächen und Abwässern, an Teichen, in Klärschlammbecken und Talsperren, wenn deren Wasserstand im Sommer gefallen ist. Voraussetzung ist, daß Absätze – und seien es dünne Häute – von stickstoffhaltigem Schlamm zurückbleiben, auf denen sich die Bidentetea-Arten als Wärmekeimer und wahrscheinliche Nitrophyten entwickeln können. Wenn auch mit Bidens frondosa, Bidens connata und der Ahnensippe der erst in Europa neu entstandenen Ufer-Spitzklette Xanthium albinum Neophyten aus Nordamerika am Aufbau beteiligt sind, so ist doch am mitteleuropäischen Indigenat der meisten Arten nicht zu zweifeln. Ähnlich geartete Standorte dürften in der Naturlandschaft nicht gefehlt haben; besonders um Wildtränken muß der Tritt der Tiere geeignete offene Stellen geschaffen haben. Daß der Höhepunkt der Entfaltung der Bidentetea jedoch heute bei uns überschritten ist, läßt sich aus den genannten Standortsbedingungen folgern.
Charakteristische Arten: Bidens frondosa, Bidens tripartita, Polygonum lapathifolium, Polygonum mite, Rorippa sylvestris.
|

Klasse ISOETO-NANOJUNCETEA: Annuelle Pionier- und mehrjährige kurzlebige Zwergpflanzen-Gesellschaften auf periodisch überschwemmten nackten Böden. Im Frühling und in der Frühsommerblüte manchmal mit Isoetiden (Isoetetalia) [Agrostion pourretii, Isoetion, Cicendion, Menthion cervinae]; im Spätsommer und in der Herbstblüte ohne Isoetide (Nanocyperetalia) [Verbenion supinae, Lythrion tribracteati]. Holarktisch
Beschreibung:
Ein anderer Typus von nur kurzfristig besiedelbaren Standorten entsteht an den Rändern und am Grunde von austrocknenden Tümpeln, Altwässern und Talsperren, auch von Fischteichen, die zum Ausfischen abgelassen worden sind. Dies sind die Biotope der „wechselnassen Zwergpflanzenfluren", wie ELLENBERG (1996) sie treffend nennt, von denen viele Arten ausgesprochene, unauffällige Seltenheiten unserer Flora sind.
Von hier aus sind einige der Vertreter auf zeitweise nasse Waldwege und Ackerfurchen oder im atlantischen Gebiet sogar auf großflächig vernäßte Äcker, also offenbar ähnliche Standorte übergegangen. Was diesen gemeinsam und daher für die Klasse Isoeto-Nanojuncetea ausschlaggebend ist, muß man durch qualitativen Vergleich herausschälen, da erst wenige exakte Messungen vorliegen. Die große klimatisch, edaphisch, aber auch florengeschichtlich bedingte Mannigfaltigkeit der Gesellschaften kommt in ihrem Reichtum an gut charakterisierten Assoziationen zum Ausdruck (Zahlen dazu s. PIETSCH 1973). Die Klasse hat nach bisherigem Wissensstand ein europäisch-nordafrikanisch Zentralasiatisches Areal; es sind keine verwandten Gesellschaften aus Amerika oder Ostasien bekannt.
Gemeinsam ist diesen Standorten, daß sie durch Überflutung, Pflügen oder Tritt periodisch pflanzenfrei werden. Zeitweise stehen dann feuchter Schlamm, Ton, seltener auch Löß oder Feinsand zur Besiedlung bereit. Dieses Substrat ist, da die Poren wassererfüllt sind, jedenfalls zunächst sauerstoffarm; sofern mineralischer Stickstoff vorhanden ist, muß dieser als NH4+ vorliegen. Später trocknet der Boden ab und aus; doch auch jetzt ist er noch dicht gelagert, wie die sehr langen Sickerzeiten beweisen, die MOOR (1936) fand.
An Stellen, die sicher über Jahrzehnte oder gar Jahrhunderte hin, wenn auch nur periodisch, besiedelbar waren (z. B. flache Mulden in felsigem Gelände der Mittelmeergebirge), können dauerhaft mit den Therophyten einige Geophyten, z. B. die seltenen Isoetes-Arten I. histrix und I. durieui, oder Hemikryptophyten leben. Wo abgestorbene Algenwatten oder Wasserpflanzenreste den Boden bedecken, wo Röhrichtarten stokken, wo auf verschlämmten Ackern das Getreide dennoch dicht steht, kommen keine Zwergbinsen hoch! Erfolgt keine weitere Störung, so können auf die Pioniere der Zwergpflanzenflur Ausdauernde folgen; doch meist wird die Pioniervegetation nach einigen Monaten ausgelöscht, weil der Wasserstand steigt, der Stoppelacker umgeworfen wird oder eine flachgründige Krume völlig austrocknet.
Im Gelände werden diese Zwergpflanzen nur selten höher als 10 cm. PIETSCH konnte jedoch experimentell zeigen, daß bei besserer Wasser- und Stickstoffversorgung jedenfalls einige von ihnen (Cyperus fuscus und Eleocharis ovata) einen halben Meter Höhe erreichen können; das dürfte aber kaum für alle Arten gelten. Die ökologische Eigenart der Isoeto-Nanojuncetea-Arten liegt also darin, daß sie auch dann, wenn sie nur zwergwüchsig sind, sei dies genetisch bedingt oder ökologisch erzwungen, schon nach 1-2 Monaten zur Fortpflanzung gelangen (ob alle?). Doch auch diese Fähigkeit hat ihre Grenzen; bei intensivierter Bewirtschaftung unter Verzicht auf jegliche Pause — etwa bei der Teichbewirtschaftung — haben viele Gesellschaften ihren Lebensraum verloren.
Zwergbinsenfluren leben heutzutage ganz überwiegend an menschlich beeinflußten, wo nicht geschaffenen Standorten. In der Urlandschaft müssen mögliche Habitate — etwa Tümpel, Teiche, Altwässer — in manchen Landstrichen selten und weit voneinander entfernt gewesen sein. Wie waren diese überhaupt zu erreichen, wie neu entstandene geeignete Wuchsorte zu besiedeln? Auf den Wind als Ausbreitungsagen zu setzen, hätte für die Pflanzen ebenso wenig Erfolg gebracht wie dem heutigen Naturschützer. Denn wenn die Diasporen auch klein und zahlreich sind, widerspricht dem schon der geringe Abstand vom Boden und ebenso das „Gießkannenprinzip" der Anemochorie; sie wirkt zufällig und kleinräumig. Nachweislich transportieren jedoch Wat- und Wasservögel die Diasporen mit angetrocknetem Schlamm und mit Schleimhüllen, wodurch sie an Füßen und Gefieder hängen bleiben (Epizoochorie).
Eine überlebenswichtige Eigenschaft vieler (aller?) Arten der Zwergbinsenfluren ist es weiter, eine über Jahrzehnte lebensfähige Boden-Samenbank aufzubauen. So können verschollene Seltenheiten plötzlich an Stellen mit Bodenstörung „auftauchen" und so den falschen Eindruck von junger Einwanderung oder potentieller Allgegenwart erwecken (s. dazu KRAUSE 1979, POSCHLOD 1993).
Charakteristische Arten: Agrostis pourretii, Centaurium maritimum, Cicendia filiformis, Holcus gayanus, Hypericum humifusum, Illecebrum verticillatum, Gnaphalium luteoalbum, Gnaphalium uliginosum, Juncus bufonius, Juncus capitatus, Juncus tenageia subsp. tenageia, Juncus tenageia subsp. perpusillus, Lythrum portula, Mentha pulegium, Molineriella Iaevis, Pulicatia paludosa, Radiola linoides, Scirpus setaceus, Sedum maireanum. Bryophyten: Anthoceros caucasicus, Archidium alternifolium, Bryum alpinum, Fossombronia pusilla, Pohlia camptotrachela.
|
IIb. VEGETATION OF LAKES, SPRINGS, FENS AND BOGS
|

Klasse ISOETO-LITTORELLETEA: Helophytische amphibische oligotrophe Zwergemeinschaften an Ufern dystropher Seen, nährstoffarmen stehenden oder langsam fließenden Gewässern (Littorelletalia) [amphibisches nährstoffarmes tiefstehendes Wasser (Littorellion uniflorae), Atlantische und mediterrane West-Iberische flache meso-oligotrophe Gewässer (Hyperico elodis-Sparganion), kontinentales temporäres Wasser auf lehmigen Böden (Eleocharition acicularis)]. Holarktisch
Beschreibung:
Diese Klasse hat mit der Klasse Utricularietea intermedio-minoris der Kleinwasserschlauch-Gesellschaften die Bindung an oligo- bis mesotrophe Standorte gemeinsam; im Gegensatz zu jener leben ihre Gesellschaften jedoch zum größten Teil amphibisch im Litoral der Seen und — seltener — der Teiche auf kiesigen, sandigen oder schlammigen Böden ohne Sapropel; einige Arten stoßen in den Tiefwasserbereich des Sublitorals vor. Diese andersartige Lebensweise prägt sich auch in der Dominanz einer anderen Lebensform aus: Gerade die bezeichnenden Sippen lsoetes, Littorella Subularia, Eriocaulon, Pilularia, Lobelia und auch verschiedene Juncus-Arten sind vegetativ einander ähnliche, niedrig bleibende, schmalblättrige, aerenchymreiche Rosettenpflanzen, die als lsoetiden zusammengefaßt werden können (DEN HARTOG & SEGAL 1964) (Abb. 11). Als Klassencharakterarten können in Europa Juncus bulbosus, Littorella uniflora, regional auch Eleocharis acicularis, Ranunculus reptans und wenige andere gelten.
Der Standort ist für Kormophyten ein Grenzstandort; dies mag das wissenschaftliche Interesse an ihrer Ökophysiologie bewirken (s. Tab. 11). Der Besitz einer Mykorrhiza ist für Wasserpflanzen ungewöhnlich und hier als Anpassung an den nährstoffarmen Boden einleuchtend. Eine Sedimentation – und damit ein erhöhtes Nährstoff-Angebot – ist für Isoetiden verderblich, weil sie diese nicht durch Streckung ihrer Internodien ausgleichen können. Sie nutzen intensiv den hohen CO 2-Gehalt des Wassers in den Bodenhohlräumen; dieses lnterstitialwasser kann das 5 –100fache an CO 2 enthalten und zu 85 –100 % der CO 2-Aufnahme der untersuchten lsoetiden beitragen; dabei ist die kurze Entfernung zwischen Wurzeln und Blättern sicher günstig. Erstaunlich ist der Besitz des – an sich für xerotherme Sukkulente, also geradezu ihre ökologischen Gegenspieler typischen – CAM-Syndrom; aber wie bei jenen ist ein Speicher für nächtliche Atmungskohlensäure innerhalb der Pflanze als Anpassung verständlich, findet doch im oligotrophen Milieu keine starke nächtliche CO2-Anreicherung durch Atmung statt.
Der Arealschwerpunkt der Littorelletea in Europa liegt im atlantisch-subatlantischen Gebiet des borealen und temperaten Gürtels. Ursachen dafür sind zu suchen in den dort häufigen armen Gesteinen und Böden sowie in den im Mittel weniger hohen Sättigungsdefiziten, denen die Pflanzen nach Trockenfallen ausgesetzt sind.
Sehr nahestehende Gesellschaften gibt es in Nordamerika; Tab. 12 zeigt eindrücklich die floristisch-systematische Verwandtschaft. Ranunculus reptans, Eleocharis acicularis und lsoetes setacea (ssp. asiatica) kommen auch in Ostasien vor. So zeichnet sich eine holarktische, circumpolar-disjunkt verbreitete Klasse mit mehreren, den Kontinenten eigenen Ordnungen ab.
Zu den Strandlings-Gesellschaften gehören überwiegend natürliche Gesellschaften, die über Jahrhunderte hin erhalten bleiben, wenn nicht exogene Störungen eintreten, wobei heute die Eutrophierung und damit die Förderung produktionskräftigerer Konkurrenten des Röhrichts oder der Schwimmblatt- und Laichkraut-Gesellschaften (Klasse 7) an erster Stelle steht. Da alle rd. 20 bisher aus Europa beschriebenen Assoziationen (DIERSSEN 1975, PIETSCH 1977) recht oder sogar sehr selten, alle standörtlich gefährdet, alle wirtschaftlich unbedeutend, viele aber von hohem wissenschaftlichem Wert sind, stellt sich die Frage nach ihrer Erhaltung mit besonderer Dringlichkeit (s. WITTIG 1980)
Charakteristische Arten: Antinoria agrostidea, Baldellia alpestris, Baldellia ranunculoides, Hypericum elodes, Juncus bulbosus, Juncus heterophyllus, Potamogeton polygonifolius, Ranunculus ololeucos, Sparganium angustifolium.
|

Klasse MONTIO-CARDAMINETEA: Kaltquellen-Gemeinschaften dominiert von Bryophyten und immergrünen weichen helophytischen Phanerogamen (Montio-Cardaminetalia) [oro- und kryorotemperate kalte Quellen basenarmen Wassers (Cardamino-Montion), meso- und supratemperat meist Phanerogamen in weichem Wasser (Caricion remotae), kalkhaltige Quellen, dominiert von Bryophyten (Palustriellion commutatae), oroiberisch Carpetan-Leonesische und westlich orokantabrische oligotrophe Kaltquellen (Myosotidion stoloniferae), schlammig langsam fließende kalte Quellen, dominiert von verwurzelten stolonigen helophytischen Hahnenfußgewächsen (Ranunculion omiophyllo-hederacei)]. Holarktisch
Beschreibung:
Die Gesellschaften leben im unmittelbaren Quellbereich auf überrieseltem Gestein oder Rohboden; hier zeichnet sich das Wasser aus durch Sauerstoffreichtum und durch relativ und gleichmäßig niedrige Temperaturen, verglichen mit denen anderer nasser Standorte und quellferner Gewässer. Eindrucksvoll belegen Messungen von DAHL (1956) aus Epilobion-Gesellschaften Südnorwegens die Kaltstenothermie; er erhielt in den quellnächsten Moosrasen, denen von Pohlia wahlenbergii, Sommerwerte zwischen 1,4 und 5,0 °C, Winterwerte, unter einer Schneedecke gemessen, zwischen 1,8 und 2,2 °C! Die Isothermen folgten dabei sehr gut den Grenzen der dominierenden Moosarten. Vermutlich hängen hiermit auch die Armut an Kormophyten und der Reichtum an Moosen zusammen, konnten BOGENRIEDER & WERNER (1979) doch die Empfindlichkeit selbst des „Kaltwasserspezialisten" Carex frigida gegen tiefe Temperaturen im Wurzelbereich nachweisen. Schon bei 10 °C Wassertemperatur in Hydrokultur betrug die Wuchsleistung bei gutem Nährstoffangebot nur noch 7,6 % von der bei 20 °C erzielten. In einigen wenigen Gefäßpflanzengattungen hat sich offenbar, nachdem die Anpassung gelungen war, eine weitere Artbildung abgespielt, so bei Saxifraga und Epilobium.
Die Zäsur zwischen den Quellfluren der Silikat- und der Kalkgesteinsgebiete ist so ausgeprägt, daß die Verbände vielleicht besser zu Ordnungen aufgestuft würden. Im Cratoneurion schaffen die Pflanzen selbst ihr Substrat und tragen zur Gesteinsbildung bei. Der Zusammenhang ist folgender: Das aus Kalkfelsspalten austretende Wasser ist kühl, reich an CO2 und gesättigt mit schwerlöslichem CaCO und leicht löslichem Ca(HCO3)2; das Gleichgewicht wird durch die Formel CaCO3 + CO2 + H20 Ca(HCO3)2 qualitativ wiedergegeben. An der Erdoberfläche sinkt die Konzentration an CO2 ab, weil a) der Partialdruck geringer ist als im Berginneren, b) die Wassertemperatur im Sommer steigt und damit die Löslichkeit des Gases sinkt, c) an pflanzenbewachsenen Stellen dem Quellwasser CO2 zur Photosynthese entzogen wird. Die Folge der Konzentrationsänderung ist Zerfall des Ca(HCO3)2 und damit Übersättigung mit CaCO3, welches sich als Quelltuff oder Travertin an toten und lebenden Gegenständen niederschlägt. Das tote organische Material (Blätter, Äste, Moose) wird im Laufe von Monaten zersetzt. Wenn die umgebenden Krusten erhärtet sind, stellen sie Abdrücke dieser organischen „Matrizen" dar. Sie sind zu Fossilien geworden und erlauben unter glücklichen Umständen noch nach Jahrmillionen Rückschlüsse auf die Tuffbildner und die Vegetation der Umgebung
Charakteristische Arten: Cardamine flexuosa, Carex remota, Chrysosplenium oppositifolium, Epilobium anagallidifolium, Epilobium anagallidifolium x palustre, Epilobium obscurum, Festuca rivularis, Hypericum androsaemum, Montia anmoritana, Myosotis stolonifera, Ranunculus omiophyllus, Sagina saginoides, Saxifraga clusii subsp. lepismigena, Saxifraga stellaris, Sibthorpia europaea, Stellaria alsine, Veronica serpyllifolia subsp. langei, Bryophytes (a.o.): Brachythecium rivulare, Chiloscyphus polyanthos, Jungermannia sphaerocarpa, Philonotis caespitosa, Philonotis fontana, Philonotis seriata, Philonotis tomentella.
|

Klasse OXYCOCCO-SPHAGNETEA: Decken-Moore und torfig ombrogene oder aquatische Sphagnum- und Heidegemeinschaften (Erico tetralicis-Sphagnetalia papillosi) [hyperozeanisch meist ombrotrophischer kantabrischer Atlantik (Erico mackaianae-Sphagnion papillosi), kalt ozeanische und vor dem Austrocknen resistenten nasse Torfheiden, atlantikweit (Ericion tetralicis)]. Kosmopolitisch.
Beschreibung:
Es sind keine Arten bekannt, die ausschließlich an Hochmoore gebunden wären, wohl aber solche, die dort ihren Verbreitungsschwerpunkt besitzen: es sind die Charakterarten der Oxycocco-Sphagnetea. Niedermoore dagegen haben eine Fülle eigener Pflanzenarten aufzuweisen, so fast alle Kennarten der Scheuchzerio-Caricetea. Diese klingen jeweils im Grenzgürtel zu den ombrotrophen Moorteilen hin aus, die Linie ihres äußersten Vorkommens wird exclusive fen plant limit oder Mineralbodenwasserzeiger-Grenze genannt. Schon ehe sie, vom Moorrand her, erreicht ist, sind nur noch wenige der anspruchsvolleren Niedermoorarten in die physiognomisch bereits hochmoorartige Pflanzendecke eingesprengt (z. B. Carex lasiocarpa, Potentilla palustris, Menyanthes trifoliata, Molinia coerulea); solche Moorteile pflegt man ebenfalls häufig in nicht ganz korrekter Weise als Hochmoor zu bezeichnen; besser ist es, hier von Übergangsmooren (nicht Zwischenmooren, einem obsoletenn Begriff und Ausdruck) zu sprechen. Da für die syntaxonomische Gliederung nicht einzelne Arten maßgebend sind, sondern die gesamte Artenverbindung entscheidet, entspricht die begriffliche und reale Grenze von Oxycocco-Sphagnetea gegen Caricetea nigrae nicht exakt der von Hoch- gegen Niedermoor, sondern sie ist etwas ins Niedermoor verschoben; die ein wenig nährstoffreicheren Partien erscheinen dann als Subassoziationen und Varianten.
Die ökologische Gruppe der Mineralbodenwasserzeiger ist am besten für Schweden ausgearbeitet; JENSEN (1961) hat sie für den Harz geprüft und dort bestätigen können. Doch ist sicher, daß man zwischen absolut und nur gebietsweise gültigen Mineralbodenwasserzeigern (z. B. Eriophorum angustifolium in Südskandinavien) zu unterscheiden hat. Dieser Befund weist bereits darauf hin, daß in der Natur weitere Faktoren eine vegetationsbestimmende Rolle spielen, z. B. Staubzufuhr. Gerade die feine Ansprache von Moorpflanzen auf Nährstoffzufuhr läßt erwarten, daß man heutzutage die Folgen von unbezweifelbaren Stickstoff-Immissionen fände; solche Änderungen ließen sich indessen bisher nicht nachweisen (s. z. B. JENSEN 1987). Aber auch mitten im ombrotrophen Bereich können entlang von Rüllen, in denen überschüssiges Wasser rasch abfließt, anspruchsvolle Arten leben; vermutlich werden hier in stärkerem Maße Ionen abgefangen.
Außer dem Nährstoffhaushalt weicht auch der Strahlungshaushalt des Biotops Hochmoor vom üblichen ab: Bei Einstrahlung erhitzt sich dunkler, vor allem trockener Torf stark; nachts ist die Ausstrahlung und damit Abkühlung besonders hoch; dazu kann seitliche Kaltluft auch in Wald-umstandene Moore einfließen. Selbst im August maß HÖLZER (1977) z. B. in einem Schwarzwälder Moor in 1000 m Seehöhe frühmorgens 0 °C, am Mittag darauf aber 45 °C. Im Juli gab es Reif und vereiste Schlenken
Charakteristische Arten:
Alisma lanceolatum, Apium nodiflorum, Carex elata subsp. reuteriana, Cyperus eragrostis, Eleocharis palustris, Galium palustre, Glyceria declinata, Holcus mollis subsp. reuteri, lris pseudacorus, Lycopus europaeus, Lythrum salicaria, Oenanthe crocata, Phragmites australis, Rorippa nasturtium-aquaticum, Typha angustifolia, Typha latifolia, Veronica anagallis-aquatica, Veronica linkiana.
|

Klasse PHRAGMITO-MAGNOCARICETEA: Moorige, sumpfige, lakustrische und flussartige Helophytengemeinschaften (Helostadion) dominiert von mehrjährigen Graminoiden, Seggen, Stauden und Kräutern an Süss- und Brackgewässern. Hohe Süsswasser Graminoide (Phragmitetalia) [nur eine Allianz (Phragmition australis: Phragmitenion australis, großes Graminoid - empfindlich für zeitweilige Drainage, Bolboschoenenion maritimi, mittelgroßes Graminoid - resistent gegen Austrocknung; gekennzeichnet durch Bolboschoenus maritimus subsp. maritimus und Scirpus triqueter)]; Große Süßwasser-Seggen (Magnocaricetalia) [Sümpfe, Seen und Ufer, eurosibirisch und mediterran (Magnocaricion elatae), Flüsse, mediterrran west-iberisch (Caricion reuterianae), Madeira (Deschampsion argenteae)]; Pioniere, kurze rhizomatöse Kräuter und Graminoide von Süßwasserströmen und Pools mit wechselndem Wasserstand (Nasturtio-Glycerietalia) [hohes steifes Graminoid (Glycerio-Sparganion: Glycerio-Sparganienion, hoher Wasserstand, mit nur gelegentlicher Austrocknung, Glycerienion Fluitantis-Pools und langsam fließende flache Gewässer. Mit Austrocknungsperioden, Phalaridenion arundinaceae-Bäche mit langen Drainage-Perioden, die von Phalaris arundinacea überwiegend auf reichen Böden und Oenanthe crocata auf armen Böden charakterisiert sind), kriechend in nährstoffreichen Gewässern (Rorippion nasturtii-aquatici)]; Seggen und Graminoide in Brackwasser (Bolboschoenetalia compacti: Bolboschoenion compacti). Kosmopolitisch, aber fehlend in polaren und verwandten sehr kalten Bioklimaten.
Beschreibung:
Sie ist in der ganzen Holarktis verbreitet. In anderen Florenreichen leben ähnliche Gesellschaften, die aus nah verwandten Arten aufgebaut werden; wahrscheinlich handelt es sich also um eine kosmopolitische Klassengruppe.
Die floristische und standörtliche Großgliederung ist im BS 10 wiedergegeben. Zahlreiche Kennarten höherer Syntaxa erreichen in niedrigen Einheiten Maxima an Menge und Stetigkeit und gelten daher für diese ebenfalls als Kennarten, wenn auch eben
mit geringerer Bindung.
Die Klasse bietet ein gutes Beispiel dafür, daß zunächst verwirrende Unterschiede in den Systemen auf unterschiedliche Einstufung bestimmter „Blöcke" von eindeutig nahe „verwandten" Gesellschaften zurückzuführen und damit durchschaubar sind. So wird z. B. die Klasse Bolboschoenetea (Brackwasser- und Tideröhrichte) nur noch als Verband innerhalb der Phragmitetalia geführt. Ebenso läßt sich die Grenze zwischen den eu- und den oligotraphenten Seggensümpfen verschieden ziehen.
Die ökologischen Unterschiede zwischen den 4 Verbänden: Ein Vegetationsprofil von einer freien Seefläche zum höher gelegenen Ufergelände veranschaulicht qualitativ den Unterschied zwischen Röhrichtverbänden und Magnocaricion: Höheres Niveau über dem See- und Grundwasserspiegel bewirkt geringere Überflutung; damit trocknen die Magnocaricion-Bestände zeitweilig oberflächlich ab, ihr Oberboden ist besser durchlüftet (gleiche Bodenart vorausgesetzt).
Quantitativ sind die Verhältnisse nicht so klar: Die Grenze zwischen den Verbänden liegt in den verschiedenen Regionen nämlich nicht in der gleichen Höhe über dem Grundwasserspiegel, weil der Wasserhaushalt zusätzlich durch die oft abweichenden Bodenarten und Niederschlagsmengen bestimmt wird; die gesamte Wasserbilanz abe entscheidet über die Lebensmöglichkeit der Pflanze, nicht ein einzelner Standortsfaktor.
Die Sonderung des Bolboschoenion beruht auf dessen Halophytismus. Bei den Verbänden 3 und 4 spielt der Nährstoffgehalt innerhalb der Assoziationen eine differenzierende Rolle; er ist oft deutlicher in qualitativen Stufen aus den Trennarten abzuleiten als aus (notwendigerweise zeitlich und räumlich punktuellen) Messungen.
Charakteristische Arten:
Alisma lanceolatum, Apium nodiflorum, Carex elata subsp. reuteriana, Cyperus eragrostis, Eleocharis palustris, Galium palustre, Glyceria declinata, Holcus mollis subsp. reuteri, lris pseudacorus, Lycopus europaeus. Lythrum salicaria, Oenanthe crocata, Phragmites australis, Rorippa nasturtium-aquaticum, Typha angustifolia, Typha latifolia, Veronica anagallis-aquatica, Veronica linkiana.
|

Klasse SCHEUCHZERIO-CARICETEA FUSCAE: Sumpf-, Fenn- und Moorgemeinschaften von kleinen Seggen und Bryophyten Holarktischer Kaltgebiete. Pioniere im oligo-dystrophischen Torfmoos (Scheuchzerietalia palustris: Rhynchosporion albae), oligotropher Torf und mineralisch torfige Böden (Caricetalia nigrae) [supra und orotemperate und verwandte Gebirgsklimata (Caricion nigrae), oro und cryoromediterranes Nevadensisches Klima (Festucion frigidae), Atlantisch thermo- und mesotemperates und verwandte ozeanische Klimata (Anagallido-Juncion bulbosi)]; (Caricetalia davallianae) [Eurosibisches und mediterranes meist in großen Höhen (Caricion davallianae), Pionier basophil Arktisch und Alpin (Caricion maritimae)]. Holarktisch
Beschreibung:
Sümpfe und Niedermoore (= Flachmoore = Kleinseggenmoore) zeichnen sich dadurch aus, daß Grund-, Quell- oder Sickerwasser den Boden langfristig durchtränkt; sie trocknen nur ganz oberflächlich ab. In Mooren kommt es dabei zu einer Hemmung der Streuzersetzung, in Sümpfen nicht; Moore bilden daher Torf als semiterrestrische Humusform, d. h. die Struktur der Bestandteile ist mikroskopisch noch erkennbar; bei Sumpfhumus ist dies nicht der Fall. Auf Torfbildung und -wachstum sei erst bei den Hochmooren näher eingegangen (s. S. 256), wo diese Prozesse den Standort stärker bestimmen als bei den Niedermooren. Diese stehen den Sümpfen noch so nahe, daß manche Gesellschaften an beiden Standorten vorkommen; wollte man das in den deutschen Namen zum Ausdruck bringen, so würden diese zu schwerfällig. Im Alpenvorland lassen sich aufgrund von Differentialarten (Sesleria varia, Gentiana verna Pinguicula alpina u. a.) sogar innerhalb der Kalkflachmoore diejenigen Bestände, deren Wurzelboden mineralreich ist, von denen auf reinem Torf unterscheiden.
Die beiden Niedermoor-Ordnungen leben in Quellmulden, an durchsickerten Hängen, an Seen oberhalb des Großseggengürtels, auch am Rande von Hochmooren, wo sie dann mit Oxycocco-Sphagnetea-Gesellschaften in Kontakt treten und entsprechende Gradienten bilden. Die Schlenkengesellschaften, klarer: Scheuchzerietalia sind von ihnen ökologisch abgesetzt durch stehendes, nicht sickerndes Wasser; dies führt in flachen Schlenken u. a. zu Temperaturextremen; solche kommen auch in oligotrophen Hochmoorgewässern vor.
Insgesamt sind die Nährstoffverhältnisse der Scheuchzerio-Caricetea-Gesellschaften trotz gelegentlicher leichter Überschneidungen in einzelnen Faktoren deutlich günstiger als die der Oxycocco-Sphagnetea.
Charakteristische Arten:
Agrostis canina, Anagallis tenella. Arnica montana subsp. atlantica,. Carex demissa, Carex echinata, Carex nigra, Epilobium palustre, Eriophorum angustifolium, Juncus cf. alpinoarticulatus, Juncus articulatus, Menyanthes trifoliata, Ranunculus flammula, Scutellaria minor, Veronica scutellata,. Viola palustris, Wahlenbergia hederacea, Bryophyten: Bryum pseudotriquetrum, Calliergon stramineum, Calliergonella cuspidata, Drepanocladus fluitans, Sphagnum auriculatum, Sphagnum cuspidatum, Sphagnum russowii, Sphagnum squarrosum. Sphagnum subsecundum s. l.
|
Annex 4 - Habitate der Seen der Serra da Estrela und ihrer Umgebung (Annex I der Habitat-Richtlinie 92/43/EWG - in Portugal in der portugiesischen Legislation ‘anexo B-I do Decreto-Lei n.º 49/200, de 24 de fevereiro’ verankert)
Das Habitat (deutsch laut Duden mit Endbetonung,[1] von lateinisch habitat ‚[es] wohnt‘, mit Anfangsbetonung), im Deutschen meist Lebensraum, bezeichnet in der Biologie den charakteristischen Aufenthaltsbereich einer bestimmten Tier- oder Pflanzenart[2] beziehungsweise den durch spezifische abiotische und biotische Faktoren bestimmten Lebensraum, an dem die Art in einem Stadium ihres Lebenszyklus lebt.[3] Der Begriff Habitat wurde ursprünglich nur autökologisch, also auf eine Art bezogen verwendet. Mittlerweile wird er auch in synökologischem Zusammenhang als Synonym für Biotop verwendet, so dass auch die Lebensstätte einer Gemeinschaft mit Habitat bezeichnet wird.[4] Dies ist vor allem auf den Einfluss aus dem englischen Sprachraum zurückzuführen. So definieren Campbell und Reece ein Habitat als einen räumlich abgrenzbaren Teilbereich eines Biotops.[5] Der Begriff wird auch in anderen Wissenschaften verwendet. In der Botanik, speziell der Vegetationsökologie (Pflanzensoziologie), wird anstelle von Habitat im autökologischen Sinne meist vom Standort gesprochen. (Wikipedia)
Folgende Habitate werden vom ICNF (Instituto de Conservação da Natureza e Florestas, Portugal) für die Seen der Serra da Estrela und ihre unmittelbare Umgebung im Rahmen der Umsetzung der FFH-Richtlinie angegeben :
Habitate 3130, 3260, (4030, 4060, 4090, 6160) 6230*, 7140, 8220
3 Süsswasserhabitate
31 Stehende Gewässer
3130 [PDF 484 KB] Águas paradas, oligotróficas a mesotróficas, com vegetação da Littorelletea uniflorae e/ou da Isoëto-Nanojuncetea
3130 Águas estagnadas, oligotróficas a mesotróficas, com vegetação da Littorelletea uniflorae e ou da Isoëto-Nanojuncetea
32 Fliessgewässer - Gewässerverläufe mit natürlicher und semi-natürlicher Dynamik (kleine, mittlere und grosse Flussbette), in denen die Wasserqualität keinen signifikativen Änderungen unterliegt
3260 [PDF 322 KB] Wasserläufe basaler und mittlerer Gebirgsgürtel mit Vegetation des Ranunculion fluitantis und des Callitricho-Batrachion
3260 Cursos de água dos pisos basal a montano com vegetação da Ranunculion fluitantis e da Callitricho-Batrachion
4 Heiden und Gebüsche der temperaten Zonen
6 Natürliche und semi-natürliche Rasenformationen
61 Natürliche Rasen
62 Seminatüriche trockene Rasenformationen und Gebüsche
Formações herbáceas secas semi-naturais e fácies arbustivas
6230* Artenreiche Borstgrasrasen (Nardus) auf silikatischen Substraten in Berggebieten (und submontanen Bereichen in Kontinentaleuropa)
6230* Formações herbáceas de Nardus, ricas em espécies, em substratos siliciosos das zonas montanas (e das zonas submontanas da Europa continental)
7 Hochmoore, Niedermoore und Sümpfe
71 Saure Sphagnum -Torfmoore
7140 Übergangs-Torfmoore und Schwingrasenmoore
7140 Turfeiras de transição e turfeiras ondulantes
8 Felsige Habitate und Höhlen
82 Felshänge und Felsspalten-Vegetation
8220 Vertentes rochosas siliciosas com vegetação casmofítica
(Fussnoten und Bibliographie: Da sich Fussnoten einschl. Bibliographie
nicht ohne weiteres aus Google-Docs heraus nach Blogger übertragen
lassen, können diese im veröffentlichten Originalartikel eingesehen
werden -> https://docs.google.com/document/d/1zpBupP4InPNty2hO59kq-02CMPuirh4aVY4B8PVDCYA/pub)