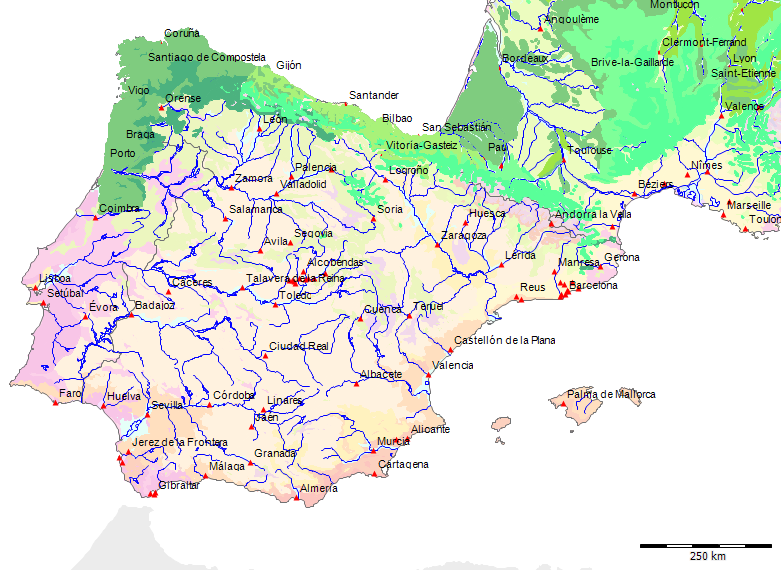
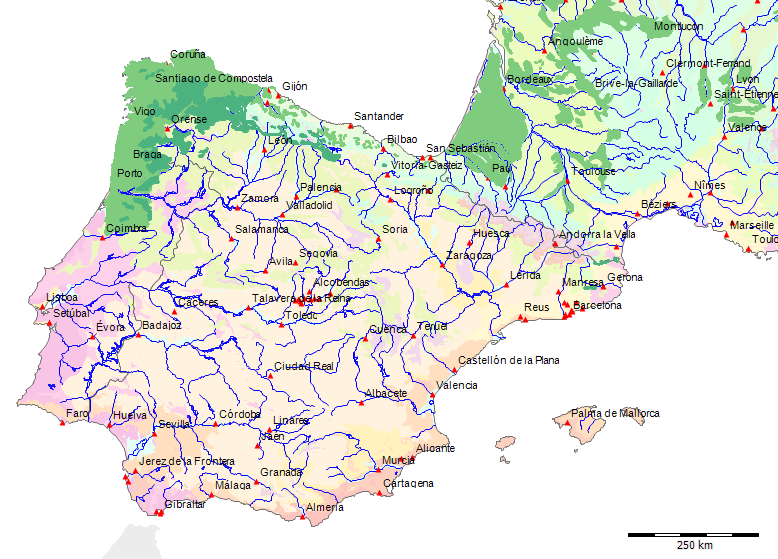
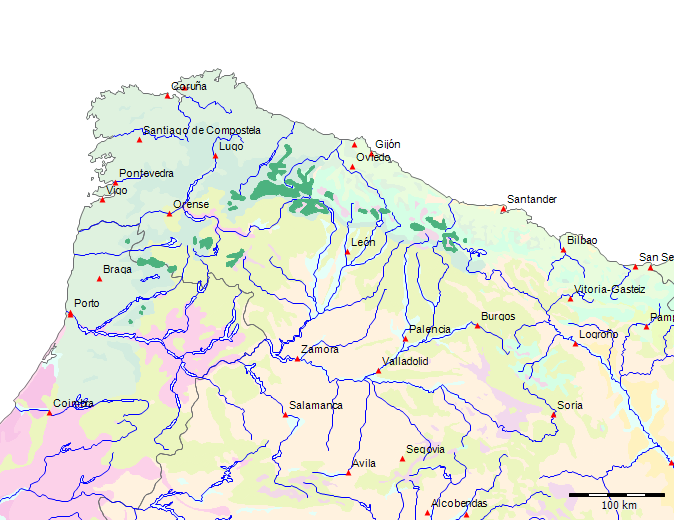
Karte der Potentiellen Vegetation[71]
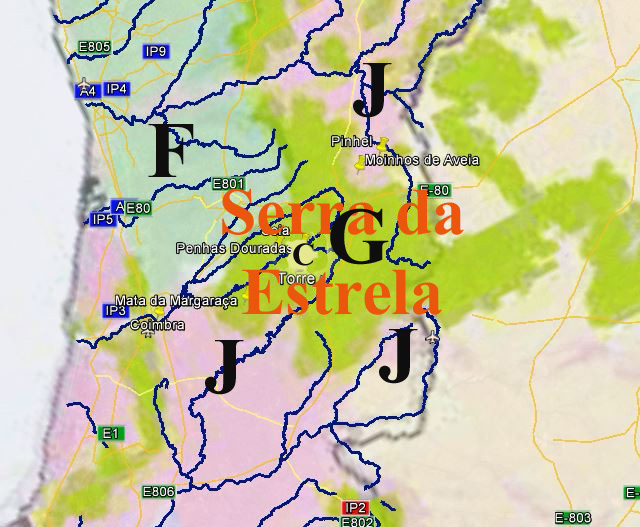 |
Potentielle Vegetation der Serra da Estrela (Nach Bohn et al. 2002)
(C - Subarktische, boreale und nemoral-montane Lichtwälder sowie subalpine und
oromediterrane Vegetation[72]
F - Mesophytische sommergrüne Laubwälder und Nadel-Laubwälder[73]
G - Thermophile sommergrüne Laubmischwälder[74]
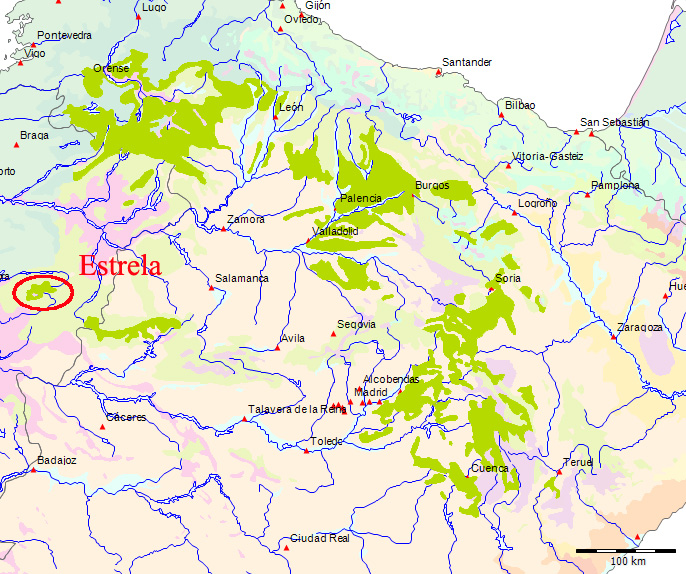
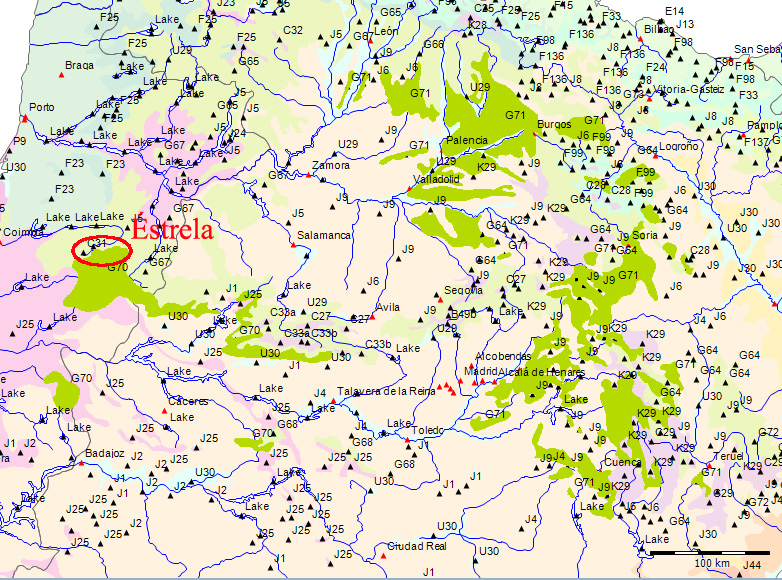
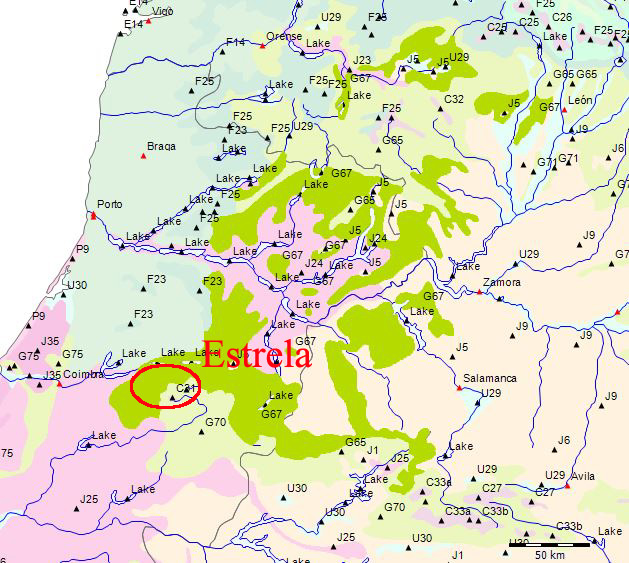
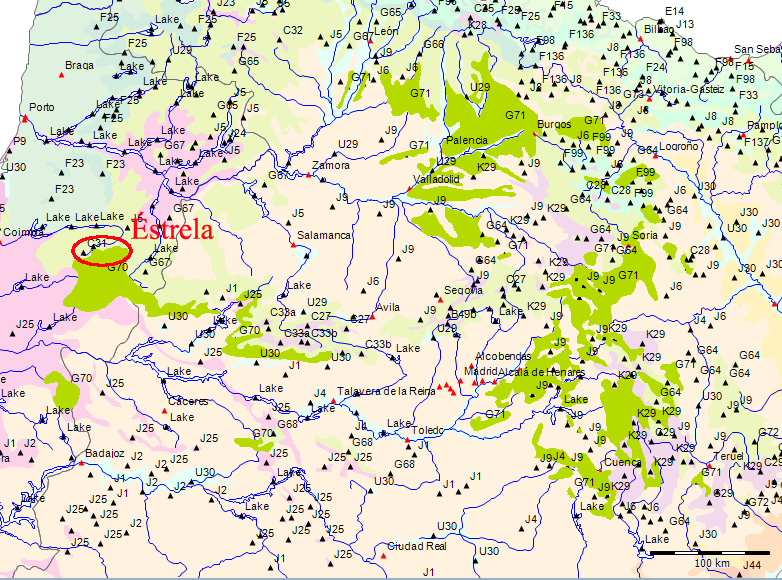
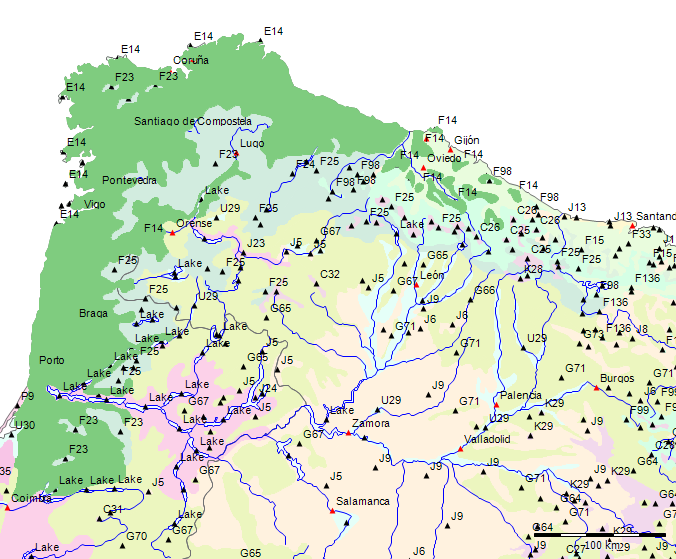
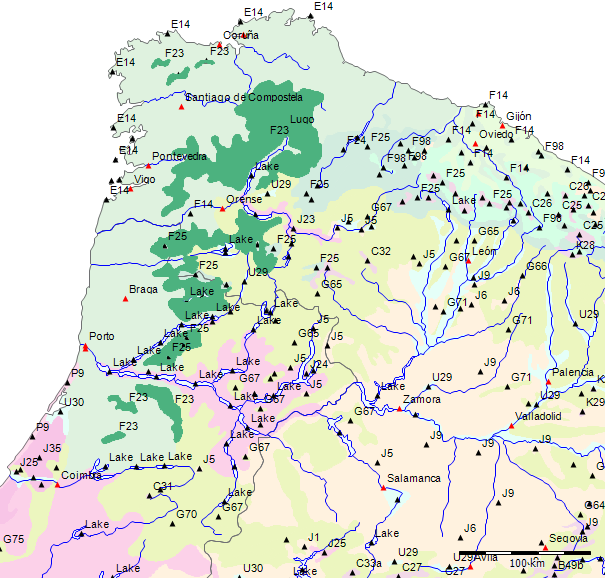
Wenn wir uns die Region der Serra da Estrela in der Karte der Potentiellen Vegetation von Europa (BOHN et al. 2002) anschauen, sehen wir, dass hier 2 Formationen für die Kernzone der Serra da Estrela kennzeichnend sind: die Formation G der “Thermophilen sommergrünen Laubwälder” und die Formation C der “Subarktischen, borealen und nemoral-montanen Lichtwälder sowie der subalpinen und oromediterranen Vegetation” mit der Unterformation C.3. (Kartierungseinheit,C31).
Am Rande der Serra da Estrela finden sich zwei weitere Formationen, - am nordwestlichen Rand die Formation F der “Mesophytischen Laubwälder und Nadel-Laubwälder”, und an den nordöstlichen und am südlichen Rändern die Formation J der “Mediterranen Hartlaubwälder und -gebüsche”.
Damit spiegelt die Karte der potentiellen Vegetation Europas die
aufgrund der Höhenzonierung vorgenommene Einteilung in Bioklimatischen
Varianten von Jan JANSEN (2002) wider, der in den unteren und mittleren
Höhenstufen an der nordwestlichen Flanke die meso- und supra-temperaten Gürtel, in den unteren und mittleren Höhenstufen der südöstlichen Flanke die meso- und supra-mediterranen Gürtel, unterscheidet. Für die temperaten und mediterranen Gürtel der unteren und mittleren Höhenstufen wird bei BOHN et. al. zwar
nur eine einzige potentielle Formation (G) thermophiler sommergrüner
Laubwälder postuliert, aber die in den Bioklimatischen Varianten
vorgenommenen Unterscheidungen zwischen temperaten Klimavarianten im
Nordwesten und und mediterranen im Südosten des Gebirges werden durch
die an den Rändern der Serra da Estrela angrenzenden borealen bzw.
mediterranen Formationen in der Karte von Bohn ebenfalls angezeigt. Auch
die oro-temperate (alpine) Stufe findet in der Karteneinheit (KE) C31
als oro-mediterrane (subalpine) Stufe teilweise eine Entsprechung. Die
unterschiedliche Einstufung der hohen Stufe, einmal als oro-temperat,
zum anderen als oro-mediterran, hängt zum einen mit der Uneinigkeit der
Experten bzgl. der Einstufung der Serra da Estrela als dem mediterranen
oder dem borealen Pflanzenreich zugehörig zusammen (siehe Fußnote 49
dieser Arbeit), zum anderen aber auch damit, dass Uneinigkeit darüber
besteht, ob das Hochplateau der Serra da Estrela über oder unter der
Baumgrenze liegt, und somit als alpine oder subalpine Höhenzone
anzusehen ist.
Der
Karte der Potentiellen Vegetation Europas erlaubt es, die potentielle
Verbreitung und die Bedeutung von Vegetationstypen und -mustern nicht
nur für begrenzte Gebiete wie das der Serra da Estrela, sondern für die
gesamte Iberische Halbinsel und den gesamteuropäischen Raum zu
beurteilen. Wir schauen uns deshalb im Folgenden den Bezug der für die
Estrela angenommenen potentiellen Vegetationsmuster zum Gesamtareal der
Iberischen Halbinsel an. Ausführliche Erläuterungen und Beschreibungen
zu den in der Karte verwendeten Vegetationskategorien (Formationen,
Subformationen, Untergruppen und Kartiereinheiten) finden sich im
Handbuch in englischer[76] und deutscher[77]
Sprache. Weiterhin enthält die Datenbanksoftware zur Vegetationskarte
zusätzlich zu einer Abfragemöglichkeit und Darstellung selektierter
Kartiereinheiten übersichtliche Datenblätter einschl. Der
Literaturverweise zu den Kartiereinheiten.
Die Formationen der potentiellen Natürlichen Vegetation der Serra da Estrela
Die Formation C “Subarktische, boreale und nemoral-montane Lichtwälder sowie subalpine und oromediterrane Vegetation” ist die spektakulärste Erscheinung in der Serra da Estrela. Sie stellt mit der Kartierungseinheit C31 ein sehr begrenztes Gebiet der Hochebene der Serra da Estrela mit einem subalpinen-alpinen Klima dar. Die systematische Beschreibung der in der Karteneinheiten findet sich in dem Handbuch zur “Karte der Natürlichen Vegetation Europas”[78]. Die Formation C wird darin folgendermaßen charakterisiert (pp. 143-45)[79]:
Charakterisierung und typologische Abgrenzung; geographische Verbreitung
Diese
Formation stellt sowohl im hohen Norden als auch in den borealen,
nemoralen und mediterranen Gebirgen die äußersten Vorposten aufrechter
und mehr oder weniger geschlossener Gehölzvegetation dar. Während im
subarktisch-nordborealen Raum Birken, besonders Betula pubescens subsp. czerepanovii, und teilweise Picea obovata dominieren, treten in den mittel- und südeuropäischen Gebirgen (Pyrenäen, Alpen, Karpaten) Pinus cembra und Larix decidua bzw. die beiden Kleinarten von Pinus mugo agg. (P. uncinata und P. mugo) bestimmend in diesen Lichtwäldern auf. In den Alpen kommen Strauchgesellschaften mit Rhododendron spp., Salix spp., Alnus alnobetula hinzu. In den südeuropäischen Gebirgen von Spanien bis zum Balkan treten je nach Gebiet Pinus uncinata (Pyrenäen, SW-Alpen), P. mugo (Apennin, Balkan), P. sylvestris (Zentralspanien), P. peuce (Balkan) oder Juniperus communis subsp. alpina bestandsbildend auf. Der Kaukasus besitzt eigene Baum- und Straucharten in der subalpinen Stufe, insbesondere Betula litwinowii, Acer trautvetteri, Rhododendron caucasicum und Quercus macranthera.
Im Norden besteht ein gleitender Übergang zu den südarktischen und Strauch-Tundren sowie zu den Gebirgstundren (Unterformation B.1),
in den Alpen und den anderen Gebirgen der nemoralen Zone werden die
subalpine Strauch- und Zwergstrauchvegetation dazu gezählt, die nach
oben anschließende alpine Stufe wird dagegen von verschiedenen baum- und
strauchfreien alpinen Vegetationstypen (Unterformation B.2)
eingenommen. Für die Abgrenzung der Lichtwälder gegen die arktischen
Tundren ist das Vorkommen einer – wenn auch sehr lichten und
niedrigwüchsigen – Baumschicht kennzeichnend. Die Trennung von den
nordborealen Nadel(misch)wäldern (Formation D) ist dagegen wegen fließender Übergänge recht schwierig.
Die Hauptvorkommen der Formation C
liegen am nördlichen Rand der borealen Zone (auch als subarktische
Unterzone bezeichnet), in den Skandinavischen Gebirgen, in Island und in
der subalpinen Stufe der südlichen Hochgebirge (Pyrenäen, Alpen,
Karpaten, Balkanische Gebirge, Kaukasus) (vgl. Karte 6). Besonders
großflächig ausgebildet und fast geschlossen ist das Verbreitungsgebiet
von den Skandinavischen Gebirgen über die Halbinsel Kola bis zum Polar
Ural. In den übrigen Mittel- und Hochgebirgen – außerhalb der Pyrenäen,
Alpen und des Kaukasus – sind die Vorkommen eher kleinflächig und
isoliert.
Bestandesstruktur und Physiognomie
Lichtwälder
weisen eine drei- bis vierschichtige Struktur auf. Neben der meist
lockeren Baumschicht ist oft eine Strauchschicht vorhanden, die
Krautschicht ist häufig zwergstrauchreich und kann nochmals gestuft
sein, und die Bodenschicht setzt sich aus Moosen und Flechten zusammen.
Neben
sommergrünen Baum- und Straucharten bauen vor allem Koniferen sowie
etliche immergrüne Sträucher und Zwergsträucher (namentlich Ericaceae),
deren Anteil sich nach Süden vergrößert, die Bestände auf. In der
Krautschicht kommen neben Zwergsträuchern – insbesondere bei
ausreichender Feuchtigkeit – oft auch Schaftpflanzen vor. Lichtwälder
sind wie der Name sagt meist lockerwüchsig, weshalb ausreichend Licht zu
den Sträuchern und Zwergsträuchern im Unterwuchs gelangt, so daß auch
hier die Mehrzahl der Arten als heliophil gelten kann. Allerdings können
in der Formation auch ziemlich dichte und schattige Wälder vertreten
sein wie z. B. geschlossene Zirbenwälder in den Alpen.
Floristische Zusammensetzung (Artengefüge)
Floristisch
zeigt die Formation eine Reihe von Gemeinsamkeiten. Boreale
Florenelemente sind innerhalb Europas weit verbreitet und können in fast
allen Teilen des Areals vorkommen (z. B. Vaccinium myrtillus, V. uliginosum, Arctostaphylos uva-ursi, Empetrum nigrum, Trientalis europaea u.
a.). Während die Formation in den nördlichen Gebirgen recht einheitlich
ausgebildet ist, wird die floristische Differenzierung nach Süden immer
größer, und es treten neben Endemiten auch andere Florenelemente (z.
B. oromediterrane oder kolchische) hinzu.
Stellung im pflanzensoziologischen System (Syntaxa)
Die
soziologische Stellung der Vegetationseinheiten innerhalb der Formation
ist recht unterschiedlich, allen gemeinsam ist jedoch das Auftreten von
Pflanzengesellschaften der Klasse Vaccinio-Piceetea
Br.-Bl. 1939. In der Gruppe der subalpinen und oromediterranen
Vegetation (Untergruppe C.2 der Formation) lassen sich namentlich in der
natürlichen Begleitvegetation neben der vorher genannten noch mehrere
andere Klassen unterscheiden: Betulo-Adenostyletea Br.-Bl. et Tx. 1943, Loiseleurio-Vaccinietea Eggler ex Schubert 1960, Junipero sabinae-Pinetea Rivas-Martínez 1964, Erico-Pinetea Horvat 1959 und – je nach pflanzensoziologischer Auffassung – auch Seslerietea albicantis Oberdorfer 1978 corr. Oberdorfer 1990.
Makroklimatische Gegebenheiten
Ein
Teil der Formation gehört zur borealen Klimazone und nimmt dort die
nördlichsten bzw. höchsten Bereiche nahe der Waldgrenze ein. Die
Jahresmitteltemperatur liegt dort zwischen -1 und 2 °C, die mittleren
jährlichen Niederschläge betragen zwischen 400 mm und 2000 mm und die
Vegetationszeit ist relativ kurz. In der südlichen temperaten Zone sind
in der subalpinen Stufe der Gebirge ähnliche klimatische Bedingungen zu
finden. In den meridionalen Gebirgen steigt die Jahresmitteltemperatur
an (Spanien bis 9 °C, Apennin bis 5 °C); hier kommt jedoch neben den
winterlichen kalten Temperaturen die sommerliche kurze
Niederschlagsdepression als klimatische Besonderheit hinzu. Als
wesentlicher klimatisch differenzierender Faktor innerhalb der Formation
ist jedoch die sich von Westen nach Osten verstärkende Kontinentalität
zu sehen. Neben der hygrischen Kontinentalität (innerhalb der Gebirge
mit Sommerregenmaxima als „Trockentäler“ ausgeprägt) wirkt sich auch die
thermische Kontinentalität mit großen Temperaturschwankungen auf die
Zusammensetzung der Vegetation aus. So liegt das mittlere Januarminimum
in den ozeanischen Gebieten (Island, Westnorwegen) bei -5 °C bis 2 °C,
im kontinentalen Bereich (norwegischer und finnischer Teil von Lappland,
Rußland) meist unter -10 °C. Die vorherrschenden Baumarten müssen
deshalb vor allem in kontinentaleren Gebieten eine ausgeprägte
Frostresistenz aufweisen.
Standortbedingungen
Für
die gesamte Formation gibt es nur wenige gleichbleibende edaphische
Standortbedingungen. Als Böden treten – besonders unter Nadelwäldern –
Podsole auf. Das Geländerelief bewirkt starke Differenzierungen zwischen
felsigen Kuppen und feuchten, oft vermoorten Mulden.
Rolle im Landschaftsgefüge
Lichtwälder
formen in allen Regionen die klimabedingte Waldgrenze, einerseits zur
Tundra hin, andererseits zur alpinen Vegetation. Im subarktischen
Tiefland bilden sie die zonale Vegetation, in den Gebirgen den
Höhengürtel im Übergang von den Nadelwäldern zu den offenen Matten und
Rasen.
Erhaltungszustand, Landnutzung, Ersatzgesellschaften; Naturschutz
Lichtwälder
sind in der borealen Zone meist gut erhalten. In den südlichen Gebirgen
wurden sie vor allem durch die Nutzung für die Almwirtschaft stark
dezimiert. Hier treten als Ersatzgesellschaften verschiedene
Zwergstrauchgesellschaften, Rasen, Hochstauden- und Lägerfluren auf.
Gliederung in Untereinheiten
C.1 - Ostboreale Lichtwälder (Betula pubescens subsp. czerepanovii, Picea obovata, Pinus sylvestris)
Verbreitung von der Halbinsel Kola bis zum Ural. Nach Osten, in Richtung Ural, erhöht sich der Anteil von Picea obovata, Larix sibirica und Abies sibirica. Im Unterwuchs sind neben Vaccinium-Arten meist Betula nana und Ledum palustre vorhanden.
C.2 - Westboreale und nemoral-montane Birkenwälder (Betula pubescens s. l.), z. T. mit Kiefernwäldern (Pinus sylvestris)
Es
handelt sich um fast reine Birkenwälder mit Hauptverbreitung im
ozeanisch beeinflußten Bereich. In Skandinavien dringt auch die Kiefer
ein. In den kontinentaleren Gebieten sind die Wälder flechtenreich, im
ozeanischen Westen hingegen farn- und hochstaudenreich. Im südlichen
Skandinavien treten im Komplex der Birken- und Birken-Kiefernwäldern
auch nemorale sommergrüne Baumarten auf.
C.3
- Subalpine und oromediterrane Vegetation (Wälder, Krummholzgebüsche
und Zwergstrauch-Gesellschaften im Komplex mit Rasen und
Hochstaudenfluren)
Diese
Wälder und Gebüsche sind in den Hochgebirgen Mittel-, Süd- und
Südosteuropas (insbesondere Pyrenäen, Alpen und Kaukasus) verbreitet und
bilden dort den obersten Wald- bzw. Gehölzgürtel. Die wichtigsten
Nadelbäume und -sträucher sind Larix decidua, Pinus cembra, P. mugo, P. uncinata, P. sylvestris und Juniperus communis, außerdem spielen sommergrüne Laubgehölze (Betula spp., Salix spp., Sorbus spp., Acer spp., Rhododendron spp.) und Zwergsträucher eine wichtige Rolle.
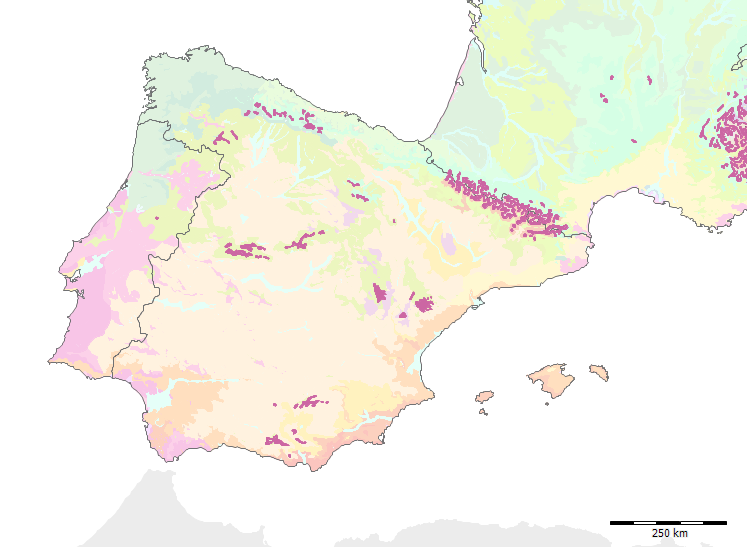
Subformation C.3 auf der Iberischen Halbinsel
|
Zur Unterformation C.3 - Iberische oromediterrane Kiefernwälder und Wacholdergebüsche (Pinus sylvestris, P. uncinata, Juniperus communis subsp. alpina) (C27-C34) (in der Estrela: die Kartierungseinheit C31) schreibt Paul Heiselmayer (pp. 161).
Im
Bereich der zentralen und südlichen Iberischen Halbinsel liegen die
Hochgebirge und ihre Gipfellagen ziemlich isoliert voneinander, so daß
sich kaum eine einheitliche Vegetation ausgebildet hat. Klimagradienten
von ozeanisch (Westen und Norden) nach kontinental (Zentralteil und
Osten) sowie von Nord nach Süd (bei zunehmender Ausprägung der
sommerlichen Trockenheit) führen zu einer weiteren Differenzierung der
Vegetation in den einzelnen Gebirgen. Die ziemlich isolierte Sierra
Nevada im Süden der Halbinsel enthält z. B. zahlreiche Endemiten. Die
oromediterranen Einheiten der Iberischen Halbinsel lassen sich in solche
mit Kiefernwäldern (Pinus sylvestris var. iberica und var. nevadensis) auf Silikat- und Karbonatgesteinen (C27, C28, C29, C30) und in solche mit Ginster-Wacholder-Gebüschen auf Silikatgesteinen (C31, C32, C33, C34) gliedern. Die weitere Unterteilung in Kartierungseinheiten erfolgt mit Hilfe geographischer Differentialarten.

Als
Natürliche Vegetation des Hochplateaus der Serra da Estrela sehen BOHN
et al. 2002) eine nord-lusitanisch oro-mediterrane Buschvegetation (Juniperus communis subsp. alpina, Cytisus oromediterraneus) mit Teucrium salviastrum, Echinospartum ibericum auf Silikatgestein an (Kartierungseinheit C31):.
C - Subarctic, boreal and nemoral-montane open woodlands as well as subalpine and oro-Mediterranean vegetation
3 - Subalpine
and oro-Mediterranean vegetation (forests, scrub and dwarf shrub
communities in combination with grasslands and tall-forb communities)
C31 - North Lusitanian oro-Mediterranean scrub (Juniperus communis subsp. alpina, Cytisus oromediterraneus) with Teucrium salviastrum, Echinospartum ibericum on siliceous rocks
Die an die Hochebene angrenzende permanente Klimaxvegetion wird nach BOHN et al. (2002) durch Pyrenäeneichenwald (Quercus pyrenaica) gebildet (Kartierungseinheit G65).
MALKUS (1985)[80] schreibt zur Vegetation der Hochebene der Serra da Estrela:
“Eine
erste floristische Bestandsaufnahme der Hochestrela wurde schon 1881
anläßlich der bereits erwähnten Expedition der "Sociedade de Geografia
de Lisboa" durchgeführt[81]:
1.
A. Henriques bestimmte 1221 Arten an Gefäßpflanzen; davon waren 0,6 %
endemisch, 7 % peninsular, 8 % mediterran, alle übrigen mittel- und
westeuropäisch.
Weitere
bedeutende Botaniker, wie Clusius, Tournefort, Daveau, Willkomm,
Ricardo da Cunha, Sampaio, Rikli u. a. arbeiteten in diesem Gebirge.
Wenngleich Henriques die gesammelten Pflanzenarten bereits 6
Höhengürteln zuordnete, so erfolgte eine der modernen Botanik Genüge
leistende begriffliche Fassung, Abgrenzung und Charakterisierung der
einzelnen Pflanzenformationen aber erst durch Braun-Blanquet et al.
(1952). Da diese Zonierung weitgehend die Grundlage für die
Habitatbeschreibung der Herpetozönosen bilden wird, bedarf es hier ihrer
ausführlichen Erläuterung.
Von
welcher Seite man sich auch dem Gebirge heute nähert, nirgends reicht
ein geschlossener Waldmantel über die 1300-1400 m Grenze, regelmäßig
liegt er tiefer. Alle Botaniker sind sich jedoch einig, daß die aktuelle
und die potentielle Waldgrenze weit auseinanderklaffen und in der Zeit,
bevor der Mensch den Wald durch Rodung, Weidewirtschaft, Köhlerei,
Streuentnahme und Brand schädigte und zerstörte, sich ein sommergrüner
Laubwald mit einzelnen immergrünen Elementen weit in die subalpine
Höhenstufe hinauf erstreckte. Darauf deuten kleine Gruppen von Betula pubescens und Einzelbäume Taxus baccata, sowie Stockausschläge an Quercus pyrenaica-Stümpfen zwischen 1400 und 1600 m hin. .
Die
von Romariz (1950) durchgeführten Pollenanalysen im moorigen
Randbereich glazialer Seen (Lagoa Comprida; Lagoa dos Covoes) in 1600 m
Höhe, zeigen uns, daß Pinus sylvestris, Betula pubescens und Taxus baccata
nach dem Rückzug der würmeiszeitlichen Gletscher die.Hochlagen des
Gebirges besiedelten. Die heute oberhalb 1550 m anzutreffenden Juniperus-Bestände (Zimbral) sind möglicherweise eine Folgeassoziation eines ursprünglichen Pinus sylvestris-Waldes.
Der Zimbral deckt beachtliche Flächen, was dem Umstand zu danken ist,
daß sich die tellerförmig flache Wuchsform des Gehölzes ebenso wie seine
sehr lückige Bestandsdichte Flächenbränden kaum Nahrung gibt. Da das
Holz selbst, im Gegensatz zu dem von Quercus pyrenaica,
keine Bedeutung für die Holzgewinnung hat und die Beeren von den Hirten
gerne zur Herstellung von Wacholderschnaps gesammelt werden, sind die
Bestände wenig gefährdet.
Die
potentielle Baumgrenze wird von einzelnen Autoren in sehr
unterschiedliche Höhenlagen versetzt: Cyren (1930) nimmt sie - mit
Sicherheit zu tief - bei 1500 man, Gaussen (1940) bei 1700 m, Brosche
(1978) bei 1800 m und Lautensach (1932) gar bei 1900 m. Die Habitatwahl
des Plateau-Endemiten Lacerta monticola
läßt den Schluß zu, daß größere Flächen bereits vor den ersten
anthropogenen Eingriffen waldfrei, bzw. mit einer sehr lückig
bestandenen Niederformation ausgestattet gewesen sein mußten;
andererseits weist das Vorkommen des Feuersalamanders in 1870 m Höhe
darauf hin, daß zumindest in geschützten Bacheinschnitten der Wald in
diese Höhen hinaufgereicht haben muß, sofern Eiselts (1958) Behauptung
richtig ist, daß das Vorkommen dieses Salamanders "immer auf eine
frühere Bewaldung hinweist, ganz gleich, ob diese durch den Menschen
oder durch klimatische Einwirkungen zum Verschwinden gebracht wurde:'
Hinsichtlich der Frage nach der potentiellen Baumgrenze neige ich daher
der Ansicht Brosches, bzw. Lautensachs zu.
Aber
selbst wenn die Waldgrenze tiefer gelegen haben sollte, ist die rezente
Vegetationsdecke des Estrelaplateaus keine natürliche, durch
Selbstregelung sich erhaltende Organismengesellschaft, sondern ein
Kunstgebilde, ein Biozönoid (sensu Schwerdfeger 1956), das nur durch
periodische Brandrodung und Beweidung am Leben erhalten wird.”
Formation G - Thermophile sommergrüne Laubmischwälder
Charakterisierung und typologische Abgrenzung; geographische Verbreitung[82]
Die thermophilen sommergrünen Laubmischwälder (Formation G)
fügen sich als überwiegend zonale Vegetation und als wechselnd breiter,
vielfach unterbrochener Gürtel zwischen die mesophilen
Fallaubmischwälder im Norden (Formation F), die Waldsteppen und Steppen im Südosten (Formation L und Formation M) und die mediterranen immergrünen Hartlaubwälder und -gebüsche (Formation J) im Süden ein (siehe Karte 13 und Übersichtskarte 1 : 10 Mio).
Ihre
größte Flächenausdehnung und Formenvielfalt erreichen sie auf der
Balkanhalbinsel und im nördlich angrenzenden pannonischen Raum. Ihr
Hauptverbreitungsgebiet liegt im Bereich der submediterranen
Florenregion (vgl. Karte 3).
Entsprechend
ihrer Übergangsstellung zwischen den Formationen F, J und L, M lassen
sie sich floristisch und standörtlich durch folgende Merkmale
charakterisieren:
- Die Baumschicht setzt sich hauptsächlich aus wärmeliebenden bzw. Trockenheitsresistenten sommer- bis wintergrünen Laubbaumarten, vorwiegend Eichen, mit submeridionalem Verbreitungsschwerpunkt zusammen (eine gewisse Ausnahme bilden die nördlichen sowie subkontinentalen Ausläufer und Vorposten mit dominierender Quercus petraea oder Q. robur).
- In der meist artenreichen Krautschicht überwiegen meso- bis eutraphente thermophile bzw. Trockenheit ertragende krautige Pflanzen, wohingegen mesophile Krautarten (im Unterschied zu den meisten Einheiten der Formation F) weitgehend fehlen. Es herrschen mitteleuropäische und submediterrane Florenelemente vor. Der Anteil mediterraner und pontischer Elemente nimmt nach Süden bzw. Südosten zu. Die nächstverwandten Vegetationseinheiten der temperaten Zone sind thermophile Eichen-Hainbuchenwälder und Waldsteppen. Im Süden gibt es fließende Übergänge zu den mediterranen Hartlaubwäldern und -gebüschen.
- Auch klimatisch nimmt das Areal der thermophilen Fallaubmischwälder eine Zwischenstellung zwischen der mitteleuropäisch-temperaten und der mediterranen sowie pontischen Region ein, wobei nach allen Richtungen fließende Übergänge bestehen. Das Temperaturklima ist wintermilder und sommerwärmer als im temperaten Bereich, jedoch winterkälter (mit regelmäßigen Frost und Schneeperioden) und nicht so extrem sommertrocken wie im eumediterranen Bereich oder in der pontisch-turanischen Steppen- und Wüstenregion.
Innerhalb der temperaten Laubmischwaldzone nehmen die Einheiten der Formation G
die trockenwärmsten Standorte ein, und sie gelten hier deshalb als
besonders thermophil, innerhalb der mediterranen Region konzentrieren
sie sich dagegen auf die niederschlagsreicheren und im Sommer besser
wasserversorgten Standorte der supramediterranen Höhenstufe. Nach oben
schließen hier vielfach thermo- bis mesophile Buchenwälder des
Mediterranbereichs an (insbesondere in Nordspanien, Süditalien und
Griechenland).
Der
größte Teil des Areals dieser Wälder liegt auf den drei großen
Mittelmeerhalbinseln: der iberischen (nördliche Hälfte), der
italienischen und der balkanischen. Auf den Mittelmeerinseln kommen der
Formation zugehörige Wälder vor allem auf Korsika, Sardinien und
Sizilien in den höheren Lagen vor, fehlen aber auf den Balearen und auf
den meisten ägäischen Inseln (mit Ausnahme von Euböa, Thasos, Samothrake
und Imroz).
Am
weitesten nach Norden dringen die thermophilen Fallaubmischwälder als
zonale Vegetation in Zentraleuropa vor: relativ großflächig in
Österreich, der Slowakei, Ungarn und Rumänien, mehr inselartig – als
extrazonale Vegetation auf Sonderstandorten – in der Schweiz, in
Deutschland, Tschechien und vor allem in Polen (besonders im zentralen
und östlichen Teil des mittelpolnischen Tieflandes). Breitere Ausläufer
nach Norden haben sie auch in Frankreich (bis zur Loire). Nach Osten
erstreckt sich das Areal dieser Wälder mit isolierten Vorkommen über die
Krimhalbinsel, die Südhänge des westlichen Kaukasus bis zu den
nordöstlichen Ausläufern des Großen Kaukasus. Im südlichen Kaukasus sind
kleinflächige Vorkommen und verwandte Einheiten in die Formation F.7
(F167-F172) integriert. Außerhalb Europas sind solche Wälder großflächig
vor allem in Kleinasien verbreitet.
In
diesem sich über 17 Breitengrade (zwischen 36º und 53º nördl. Breite)
und 57 Längengrade (zwischen 9º westl. Länge und 48º östl. Länge)
erstreckenden Areal sind die pflanzengeographischen und standörtlichen
Gegebenheiten naturgemäß sehr unterschiedlich und die natürlichen
Vegetationseinheiten entsprechend vielgestaltig.
Zum besseren Verständnis der potentiellen Vegetation der Serra da Estrela ist vor allem die Subformation G.4 der Formation G maßgeblich, da sie die potentielle Vegetation der Serra da Estrela ausser für das Hochplateau bestimmt.
G.4 Iberische supra- und mesomediterrane Quercus pyrenaica-, Q. faginea-, Q. faginea
subsp. broteroi- und Q. canariensis-Wälder
Diese
Gruppe umfaßt die supra- und mesomediterranen sommer- bis wintergrünen
Eichenwälder auf der Iberischen Halbinsel außerhalb des Areals von Quercus pubescens. In ihnen dominieren die auf das westmediterrane Gebiet beschränkten Quercus-Arten: Q. pyrenaica, Q. faginea, Q. faginea subsp. broteroi und Q. canariensis. Wichtige Mischbaumarten sind Acer monspessulanum, A. granatense und Sorbus torminalis.
Die Hauptverbreitung dieser Wälder liegt im nordiberischen Bergland und
in den dortigen Gebirgen, wo Klima und Vegetation bereits ausgesprochen
mediterranen Charakter aufweisen. Sie nehmen hier wie im Süden der
Iberischen Halbinsel die niederschlagsreicheren und kühleren Höhenstufen
ein, die im Süden nur noch relativ kleine Flächen in den Gebirgen
umfassen. Das Klima im Gebiet dieser Wälder ist dementsprechend
größtenteils mäßig warm (Jahresmitteltemperaturen 8-13 °C) und
trocken-subhumid bis humid (Jahresniederschläge 500-1000 (>1600) mm).
Die Winter sind relativ kalt (Jahresmittel des kältesten Monats -1 bis 5
°C), die Sommer warm und trocken.
Das Areal der vier Hauptbaumarten ist streng an silikatische (Quercus pyrenaica, Q. canariensis) bzw. karbonathaltige und basenreiche (Q. faginea, Q. faginea subsp. broteroi)
Gesteine gebunden, entsprechend wurde die Gliederung in die vier
Untergruppen vorgenommen. Innerhalb der Untergruppen erfolgt die weitere
Unterteilung in Kartierungseinheiten vorwiegend nach geographischen
Gesichtspunkten und mit Hilfe regionaler Trennarten (bzw. spezifischer
Regionalgesellschaften), wobei ein deutliches West-Ost- sowie
Nord-Süd-Gefälle besteht; die floristische Kennzeichnung und
Differenzierung der Gehölzbestände der einzelnen Untergruppen lässt sich
Tab. 16 entnehmen;
hinsichtlich der floristisch-ökologischen Charakteristiken der
einzelnen Kartierungseinheiten sei auf die entsprechenden Datenblätter
verwiesen.
Syntaxonomisch
werden die bodensauren (G.4.1 und G.4.4) und die basiphilen (G.4.2)
Eichenwälder verschiedenen Ordnungen und Verbänden/Unterverbänden
innerhalb der Klasse der Querco-Fagetea zugeordnet, nämlich den Quercetalia roboris, und zwar dem Quercion pyrenaicae bei G.4.1 und dem Quercion roboris bei einem Teil von G.4.4, den Quercetalia pubescentis, Quercion pubescenti-petraeae, Aceri granatensis-Quercenion fagineae bei G.4.2. Dagegen wird G.4.3 und ein Teil von G.4.4 zum Quercenion broteroi innerhalb der Klasse Quercetea ilicis gestellt.
Tab. 16 Verbreitung
der wichtigsten Baum- und Straucharten der thermophilen sommergrünen
Laubmischwälder in den Untergruppen der Formation G. (Nach BOHN et. al. 2002, pp. 328-31):

Formation G mit den Subformationen (G3, G4) auf der Iberischen Halbinsel
| |
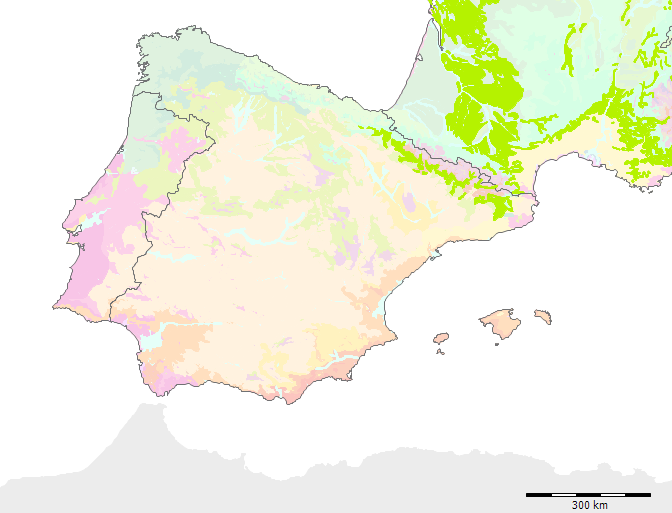
Subformation G3
G41 - Downy
oak forests (Quercus pubescens) with Buxus sempervirens, Genista
cinerea, partly Acer opalus from the south Pyrenees to the southwest
pre-Alps
| 
Subformation G4 -
mitit den Untergruppen
G4.1-G4.4 (siehe folgende Tabelle)
|
Untergruppe G4.1
G66 - North Iberian supra-Mediterranean Quercus pyrenaica-forests with Festuca heterophylla
| 
Untergruppe G4.2
G71 - North and central Iberian supra-Mediterranean Quercus faginea-forests with Cephalanthera rubra, Cephalanthera longifolia
G73 - North Iberian supra-Mediterranean Quercus faginea-forests with Spiraea hypericifolia subsp. Obovata
G74 - South Iberian supra-Mediterranean Quercus faginea-forests with Daphne laureola subsp. latifolia
|
Untergruppe G4.3
G75 - Middle Portuguese meso-Mediterranean basiphilous Quercus faginea subsp. broteroi-forests with Arisarum simorrhinum
| 
Untergruppe G4.4
G77 - Andalusian meso-Mediterranean Quercus canariensis-forests with Ruscus hypophyllum
|
Untergruppen der Subformation G4 auf der Iberischen Halbinsel
KE G65
| 
KE G67
|
KE G70
|
Kartierungseinheiten von Untergruppe G4.1, die sich mit dem Gebiet der Serra da Estrela überschneiden
G.4.1 West- und zentraliberische supra- bis mesomediterrane Quercus pyrenaica-Wälder
auf Silikatgesteinen
Diese Untergruppe besteht aus sieben Kartierungseinheiten (G64-G70) mit der sommergrünen Quercus pyrenaica
als bestandsbildender Baumart und repräsentiert bodensaure
Silikatstandorte im höheren Bergland in verschiedenen Regionen der
Iberischen Halbinsel mit Schwerpunkt in deren Nordwestteil. Die
Kartierungseinheiten stellen verschiedene ökologisch-geographische
Ausbildungen mit entsprechenden Differentialarten dar.
Soziologisch wurden diese Wälder folgenden Assoziationen innerhalb des Quercion pyrenaicae zugeordnet: Luzulo forsteri-Quercetum pyrenaicae Rivas-Martínez 1963 (G64), Holco mollis-Quercetum pyrenaicae Br.-Bl., P. Silva et Rozeira 1956 (G65), Festuco braun-blanquetii-Quercetum pyrenaicae Br.-Bl. 1967 (G66), Genisto falcatae-Quercetum pyrenaicae Penas et Diaz 1985 (G67), Sorbo torminalis-Quercetum pyrenaicae Rivas Goday ex Rivas-Martínez 1987 (G68), Adenocarpo decorticantis-Quercetum pyrenaicae Martínez-Parras et Molero 1983 (G69), Arbuto unedonis-Quercetum pyrenaicae (Rivas Goday 1960) Rivas-Martínez 1987 (G70), Cephalanthero rubrae-Quercetum pyrenaicae O. Bolòs et Vigo in O. Bolòs 1967. Im Klimaxstadium handelt es sich um mittelhohe, schattige, fast ausschließlich von Quercus pyrenaica beherrschte Wälder mit vereinzelten Mischbaumarten (Fraxinus
angustifolia, Sorbus aria, S. torminalis, Acer campestre, A.
monspessulanum, A. granatense, Quercus faginea, Q. suber, Q. ilex subsp.
rotundifolia).
Die je nach Lichtdurchlässigkeit der Baumkronen unterschiedlich
entwickelte Strauchschicht beherbergt sowohl mitteleuropäische (Crataegus monogyna, Corylus avellana, Prunus spinosa, Cytisus scoparius) wie auch submediterrane und mediterrane Arten (Berberis vulgaris, Genista florida, G. falcata), darunter auch immergrüne Sträucher und Kletterpflanzen (Arbutus unedo, Viburnum tinus, Erica arborea, E. australis, Ilex aquifolium, Hedera helix).
In der meist gut entwickelten Krautschicht kommen neben eutraphenten nemoralen Arten (Melica uniflora, Lilium martagon, Hepatica nobilis, Sanicula europaea, Viola odorata u. a.) azidophile Arten (Luzula forsteri, Holcus mollis, Teucrium scorodonia, Lathyrus linifolius, Pteridium aquilinum, Ajuga pyramidalis) und etliche südeuropäische sowie endemische Arten vor (Physospermum
cornubiense, Helleborus foetidus, Melittis melissophyllum,
Hyacinthoides hispanica, Moehringia pentandra, Paeonia coriacea u. a.).
G.4.2 Nordost- und südiberische supramediterrane basiphile Quercus faginea-Wälder
Diese Untergruppe enthält vier Kartierungseinheiten (G71, G72, G73, G74) mit der wintergrünen Quercus faginea
als bestandsbildender Baumart. Ihre Hauptverbreitung liegt im Bergland
im Nordosten der Iberischen Halbinsel, isolierte Vorkommen in den
südiberischen Gebirgen. Es handelt sich im Vergleich zu den im Nordosten
angrenzenden Quercus pubescens-Wäldern (G41)
um ausgesprochen supramediterrane Standorte mit ausgeprägter
Sommertrockenheit. Im Unterschied zu den Quercus pyrenaica-Wäldern
(G.4.1) sind die Substrate Kalksteine, Dolomite und vor allem kalkreiche
Mergel.
Soziologisch gehören diese Wälder zu folgenden Assoziationen innerhalb des Quercion pubescentipetraeae: Cephalanthero rubrae-Quercetum fagineae Rivas-Martínez in Rivas Goday et al. 1960 (G71), Violo willkommii-Quercetum fagineae Br.-Bl. et O. Bolòs 1950 (G72), Spiraeae obovatae-Quercetum fagineae O. Bolòs et Montserrat 1984 (G73), Daphno latifoliae-Aceretum granatensis Rivas-Martínez 1965.
Die zwei- bis dreischichtigen, niedrig- bis mittelwüchsigen Wälder haben in der oberen Baumschicht vorherrschend Quercus faginea mit unterschiedlicher Beimischung von Acer monspessulanum, A. granatense, A. campestre, Sorbus aria, S. torminalis, Quercus ilex subsp. rotundifolia, Q. coccifera.
Die Strauchschicht ist je nach Dichte der Baumschicht unterschiedlich
entwickelt und in der Regel artenreich. Sie enthält vor allem basiphile
submediterrane und mediterrane, meist sommergrüne Arten: neben Crataegus monogyna insbesondere Viburnum lantana, Buxus sempervirens, Amelanchier ovalis, Berberis vulgaris subsp. seroi und subsp. australis, ferner iberisch-endemische Arten wie Cytisophyllum sessilifolium[83], Genista hispanica, Spiraea hypericifolia subsp. obovata, Daphne laureola subsp. latifolia. Unter den Lianen sind vor allem Hedera helix, Lonicera etrusca und Rubia peregrina vertreten. Die Krautschicht enthält als kennzeichnende Arten Cephalanthera rubra, C. longifolia, Viola willkommii, Helleborus foetidus, Paeonia officinalis subsp. microcarpa, Primula veris subsp. columnae.
G.4.3 Portugiesische mesomediterrane basiphile Quercus faginea subsp. broteroi-Wälder
Die Untergruppe besteht aus einer Kartierungseinheit (G75), die als Arisaro simorrhini-Quercetum broteroi Br.-Bl., P. Silva et Roseira 1956 aus Portugal beschrieben und dem Verband Quercion ilicis
zugeordnet wurde. Es handelt sich um mesomediterrane Standorte auf
Kalkstein und Mergel. Die dreischichtigen Wälder bestehen in der
Baumschicht aus Quercus faginea subsp. broteroi mit Laurus nobilis und
örtlich Quercus ilex subsp. rotundifolia, in der Strauchschicht
überwiegen mediterrane, meist immergrüne Arten (Arbutus
unedo, Viburnum tinus, Daphne gnidium, Pistacia lentiscus, Quercus
coccifera, Ruscus aculeatus, Rubus ulmifolius, Osyris alba). Lianen spielen ebenfalls eine wichtige Rolle (Smilax aspera, Rubia peregrina subsp. longifolia, Asparagus aphyllus, Hedera helix). In der Krautschicht finden sich Coronilla valentina subsp. glauca, Vinca difformis, Arisarum simorrhinum, Teucrium scorodonia u. a.
G.4.4 Nordost- und südiberische mesomediterrane Quercus canariensis-Wälder auf Silikatgesteinen
Die Untergruppe enthält zwei entfernt liegende kleinflächige Kartierungseinheiten, eine in Katalonien (G76), die andere in Andalusien (G77). Es handelt sich um isolierte Vorkommen von mesomediterranen Wäldern mit der wintergrünen Quercus canariensis in relativ humiden Lagen. Die katalonischen Wälder wurden der Assoziation Carici depressae-Quercetum canariense O. Bolòs 1954, die andalusischen dem Rusco hypophylli-Quercetum canariensis Rivas-Martínez 1975 innerhalb des Verbandes Quercion ilicis zugeordnet.
Die dreischichtigen Wälder haben in der Baumschicht Quercus canariensis als bestandsbildende Art und als Mischbaumarten immergrüne Eichen (Quercus ilex, Q. suber), örtlich auch Q. faginea subsp. broteroi sowie thermo- und mesophile Fallaubbäume (Sorbus torminalis, Acer monspessulanum, Prunus avium). Die Strauchschicht besteht vorwiegend aus immergrünen mediterranen Gehölzen (Arbutus unedo, Viburnum tinus, Erica arborea, Rhododendron ponticum subsp. Baeticum, Ulex jussiaei, Ruscus hypophyllum) aber auch aus sommergrünen Sträuchern (Crataegus monogyna, Prunus spinosa). Kennzeichnende Arten der Krautschicht sind Carex depressa, Teucrium scorodonia, Gaudinia fragilis, Luzula forsteri, Polystichum setiferum sowie Pteridium aquilinum auf Schlagflächen.
Literatur
BORHIDI
1996; BRULLO, GUARINO & SIRACUSA 1998, 1999; DOING KRAFT 1955;
HORVAT, GLAVA & ELLENBERG 1974; JAKUCS 1961; OBERDORFER 1948; RAMEAU
1996b; RIVAS-MARTÍNEZ 1987; RIVAS-MARTÍNEZ, FERNÁNDEZ-GONZÁLEZ, LOIDI,
LOUSÃ & PENAS 2001 (Mskr.).
Wichtige Kartierungseinheiten der Formation G für die Serra da Estrela:
G - Thermophilous mixed deciduous broad-leaved forests
4 - Iberian supra- and meso-Mediterranean Quercus pyrenaica, Q. faginea, Q. faginea subsp. broteroi and Q. canariensis forests
4.1 - West and central Iberian supra- to meso-Mediterranean Quercus pyrenaica-forests on siliceous rocks
...
...

Die Rand-Formationen der Serra da Estrela
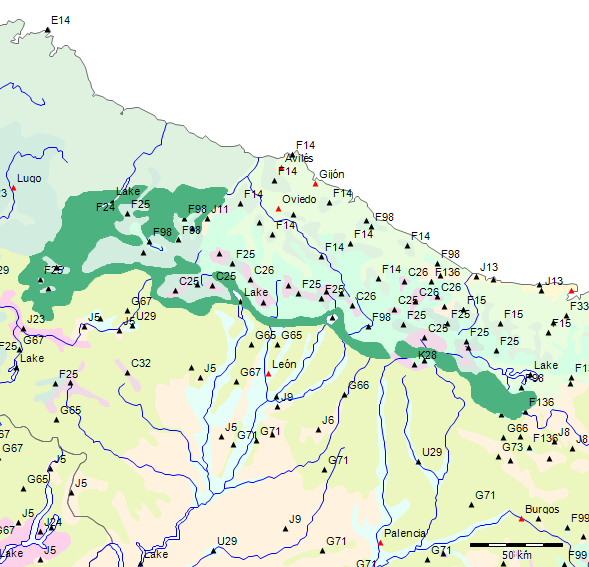
Am nordwestlichen Rand der Serra da Estrela finden sich die borealen Wälder der Formation F als potentielle Natürliche Vegetation.
Formation F
Charakterisierung der Formation F der Mesophytischen sommergrüne Laubwälder und Nadel-Laubwälder (Handbuch zur Karte der Natürlichen Vegetation Europas (p. 219) [84] ):
Charakterisierung und typologische Abgrenzung
Unter diesem Formationsbegriff (Formation F)
werden die meisten periodisch laubabwerfenden, winterkahlen
Waldgesellschaften zusammengefaßt, welche in mehr oder weniger ozeanisch
getönten Gebieten der temperaten Zone auf terrestrischen Böden zonal
vorkommen und ausschließlich oder doch vorwiegend von wenigen
Laubholzarten aufgebaut werden.
Nicht
einbezogen sind einerseits die wärmeliebenden, meist von verschiedenen
sommergrünen Eichenarten beherrschten artenreichen Laubwälder der
warm-temperaten und submediterranen Zone (Formation G), zum anderen auch
nicht die azonalen, grund- oder stauwasserbeeinflußten Gesellschaften
der Bruch-, Sumpf- und Auenwälder (Formation T und Formation U).
Zu den genannten Gruppen der sommergrünen Laubwaldgesellschaften gibt
es gleitende Übergänge, ebenso zu den Formationen der Nadelwälder (Formationen D8, D9, D12, K1
in der Übersichtskarte 1 : 10 Mio.) – hier allerdings nur im
Grenzbereich ihrer horizontalen und vertikalen Verbreitung –, ferner zur
Formation der Waldsteppen L.
Für die Kennzeichnung und Abgrenzung der Formation F gegenüber der Nadelwald-Formation D
waren floristisch-strukturelle und physiognomische Merkmale maßgebend:
Die typologische Grenzlinie fällt nämlich mit dem Umschlag zugunsten der
Nadelwaldelemente im Gesellschaftsgefüge der naturnahen Bestände
zusammen.
Die Formation F
in unserer Auffassung läßt sich mit keiner Einheit der bekannten
Formationssysteme genau identifizieren. Sie entspricht weitgehend dem
Begriff der europäischen nemoralen großblättrigen Laubwälder mancher
russischen Autoren.
Formation F mit den Subformationen (F1, F2, F5) auf der Iberischen Halbinsel
|
Subformation F1
F.1 Artenarme azidophile Eichen- und Eichenmischwälder (Quercus robur, Q. petraea, Q. pyrenaica, Pinus sylvestris, Betula pendula, B. pubescens, B. pubescens subsp. celtiberica, Castanea sativa) (BOHN et al. 2002, pp. 226)[85]
Charakterisierung und typologische Abgrenzung; geographische Verbreitung (J. Cross, J. Pallas)
Charakteristisch für bodensaure Eichen- und Eichenmischwälder ist die Vorherrschaft von Eichen, zumeist Quercus robur und Q. petraea
in der Baumschicht, sowie ein dominantes Auftreten von Azidophyten im
Unterwuchs, und zwar von Arten mitteleuropäischer oder eurasisch
temperater Verbreitung. Strauch- und Krautschicht sind meist gut
entwickelt, jedoch relativ artenarm im Vergleich zu Laubwäldern
basenreicherer Standorte oder klimatisch günstigerer Gebiete.
Das
Areal dieser Eichenwälder erstreckt sich in der temperaten Zone Europas
von der Atlantikküste bis ins westliche Rußland (Karte 9). An seiner
Südgrenze schließt es teilweise auch montane Gebiete der submeridionalen
Zone ein. Das Gesamtverbreitungsgebiet gleicht einem stark zerteilten
spitzwinkligen Dreieck, dessen Basis sich entlang der Atlantikküste von
Nordportugal (ca. 41º N) über Irland bis nach Schottland (ca. 58º N)
erstreckt. Die Spitze des Dreiecks liegt nordöstlich von Kiew (ca. 53º
N, 34º O), mit isolierten Vorposten bis nahe an die Wolga bei Kasan. Die
nördliche Grenze verläuft über Schottland, Südskandinavien, Litauen und
Weißrußland, die südliche über Nordportugal, Nordspanien,
Südfrankreich, Oberitalien, Kroatien, Bosnien-Herzegowina, Rumänien, die
Ukraine und Südrußland. Südliche Ausläufer dieser Wälder reichen bis in
die submeridionale Zone. Die Standorte weisen in der Regel durchlässige
oder aber wechselfeuchte, nährstoffarme, saure, oft sandige, z. T.
flachgründige bis felsige Böden auf. Im natürlichen Verbreitungsgebiet
der Buche – insbesondere in der Subatlantischen und Zentraleuropäischen
Provinz – werden die bodensauren Eichenwälder unter dem Konkurrenzdruck
der Buche auf wechselfeuchte Böden oder auf sehr flachgründige
Hangstandorte, die für ein Überleben der Buchenkeimlinge zu trocken
sind, zurückgedrängt. Die Beurteilung der Natürlichkeit der bodensauren
Eichenwälder ist teilweise spekulativ, denn vielerorts begünstigte die
Nieder- und Mittelwaldwirtschaft der Vergangenheit die Eiche. Die
Mehrheit der naturnahen Bestände ist daher relativ kleinflächig und
zerstreut verbreitet. Großflächige Vorkommen liegen vor allem außerhalb
des Buchenareals: in Westkantabrien und Galicien, Südwestfrankreich,
Irland und Großbritannien, Polen, Weißrußland und der Nordukraine.
Das
Areal der bodensauren Eichenmischwälder ist nicht so ausgedehnt wie das
der sommergrünen Breitlaubwälder insgesamt, denn es sind weitere
arealbegrenzende Faktoren wirksam:
- In der montanen Stufe Mitteleuropas und der submeridionalen Zone herrschen Buchenwälder vor.
- Im Norden und Nordosten des Areals ist die Konkurrenz der Nadelwälder (Formation D) der begrenzende Faktor. Auf bodensauren, nährstoffärmeren Standorten ist die Laubwaldformation im Osten Nadelbäumen eher unterlegen als auf reicheren Standorten (vgl. MONK 1966, zitiert in JÄGER 1969: 401). Daher dürften reine Laubwälder hier nur noch auf basenreicheren Standorten anzutreffen sein.
- Im Osten lassen edaphische und klimatische Faktoren (relative Niederschlagsarmut, wärmere Sommer, nährstoffreichere Böden) die Entwicklung bodensaurer Standorte nicht mehr zu. Daher verläuft die Ostgrenze der bodensauren Laubwälder weiter westlich als die Grenze der Laubwaldformation insgesamt.
- In der temperaten Zone bevorzugen bodensaure Eichenmischwälder die planare bis kolline Höhenstufe. In der submeridionalen Zone dagegen, an der Südgrenze des Areals, sind sie meist an die submontan-montane Höhenstufe gebunden. Ihre charakteristische Artenverbindung verändert sich nach Süden schrittweise, und es gelangen schließlich thermophile submediterrane Eichenwälder zur Vorherrschaft (Formation G).
Bestandesstruktur und Physiognomie (J. Cross)
Die natürliche Struktur dieser Wälder ist oft nicht eindeutig feststellbar, da sich jahrhundertelange
Beeinträchtigungen
durch den Menschen und seine Weidetiere noch heute deutlich auswirken.
Naturnahe Bestände können fünf bis sechs Schichten aufweisen, nämlich
erste und zweite Baumschicht, Strauchschicht, Zwergstrauchschicht,
Krautschicht und Moosschicht. In manchen Beständen sind jedoch nur drei
Schichten ausgebildet.
Die Baumschicht erreicht
in der Regel 60-90 % Deckung sowie eine Höhe von 15-25 (35) m. Die
Bestandeshöhe kann auf sehr flachgründigen, trockenen Böden oder in
windexponierten Lagen sogar unter 5 m bleiben, auf tiefgründigen Böden
dagegen über 30 m betragen. Die Baumschicht wird von säuretoleranten
Eichen (Quercus robur, Q. petraea) dominiert. Pionierholzarten wie Birken (Betula pendula, B. pubescens)
spielen besonders im Westen in der Regenerationsphase auf verschiedenen
Standorten eine wichtige Rolle, bleiben aber vor allem auf
wechselfeuchten Standorten auch bis ins höhere Bestandesalter in der
Baumschicht vertreten. Im Osten können Kiefer und örtlich auch Fichte
mit der Eiche die Baumschicht bilden. Deckung und Vitalität der Strauchschicht
sind abhängig von der Dichte der Baumschicht sowie von Nährstoffgehalt,
Wasserversorgung und Luftfeuchtigkeit des Standorts. Ericaceen- oder
Ginster-Arten (letztere besonders in südlicheren Gebieten) bilden
vielfach eine Zwergstrauchschicht. In den nordtemperaten Gebieten Europas verleiht die dort häufige Zwergstrauchdominanz den Wäldern ein boreales Gepräge. Die Krautschicht
wird gewöhnlich von Hemikryptophyten, namentlich von Gräsern,
Hainsimsen und Seggen aufgebaut. Geophyten sind allgemein selten, Farne
spielen jedoch in der Regel – namentlich im atlantischen und
subatlantischen Bereich – eine wichtige Rolle. Pteridium aquilinum
kann gebietsweise saisonal eine zusätzliche Schicht bilden. Bei dichtem
Bestand verdrängt die Art nahezu alle anderen Krautarten. Deckung und
Artenzahl der Moosschicht variieren ebenfalls beträchtlich. Sie erreicht
besonders im hochozeanischen Westen, wo epiphytische Gesellschaften ein
auffälliges Merkmal der Wälder sind, hohe Deckungsgrade.
Kryptogamenreiche Gesellschaften kommen jedoch auch auf dem
nordwesteuropäischen Festland auf armen und trockenen
Waldgrenzstandorten vor, wo Strauch- und Krautschicht wegen der
Konkurrenz der Baumschicht nur schlecht entwickelt sind.
Floristische Zusammensetzung (Artengefüge) (J. Pallas)
Die
bodensauren Eichenmischwälder Europas haben eine von Eichen dominierte
Baumschicht mit einem Unterwuchs, in dem azidophytische Arten
vorherrschen. Arten der temperaten Mitteleuropäischen Florenregion haben
den größten Anteil an der Artenzusammensetzung. Im Norden und Osten des
Areals nimmt der Anteil eurasisch temperater und eurasisch borealer
Sippen zu. In den südtemperaten Territorien Europas sind zahlreiche
submediterrane Arten am Gesellschaftsaufbau beteiligt, im Südwesten des
Areals sogar einige mediterrane Arten (vgl. Tab. 12). Weit verbreitete
Baumarten sind neben den beiden mitteleuropäischen Eichenarten (Quercus robur und Q. petraea) die häufig beigemischte Buche (Fagus sylvatica) sowie die eurasisch verbreiteten Arten Betula pendula und Populus tremula. Zu den häufigsten Sträuchern zählen die eurasischen Sippen Sorbus aucuparia, Frangula alnus (auf den Britischen Inseln kaum in bodensauren Eichenwäldern vorhanden) und Juniperus communis sowie – als Elemente der mitteleuropäischen Flora – Corylus avellana und in geringer Menge Crataegus monogyna. Häufige und weit verbreitete Arten der Krautschicht (s. Tab. 12) sind die Azidophyten Melampyrum
pratense, Pteridium aquilinum, Solidago virgaurea, Agrostis capillaris,
Anthoxanthum odoratum, Calluna vulgaris, Deschampsia flexuosa, Molinia
caerulea, Holcus mollis, Veronica officinalis, Carex pilulifera, Viola
riviniana, Potentilla erecta, Lathyrus linifolius und Danthonia decumbens. Die meisten azidophytischen Moose sind zirkumpolar verbreitet. Die häufigsten Arten sind Polytrichum formosum, Dicranum scoparium, Hypnum cupressiforme, Leucobryum glaucum, Pleurozium schreberi und Scleropodium purum
(Fehlstellen in iberischen Kartierungseinheiten dürften durch
unvollständige oder fehlende Erfassung der Kryptogamen in den
Vegetationsaufnahmen verursacht sein). Vertreter der submeridionalen
Zone (für Europa sind dies Arten der Submediterranen Unterregion, und –
mit weit geringerer Bedeutung – auch der Pontisch-Südsibirischen
Florenregion) sind vorwiegend in südtemperaten, insbesondere aber in
submediterranen Kartierungseinheiten anzutreffen. Zu den kennzeichnenden
Arten gehören Castanea sativa, Sorbus torminalis, Pyrus pyraster, Hieracium sabaudum und Festuca heterophylla.
In den nordtemperaten Territorien Europas fehlen im Westen die
submediterranen Arten den bodensauren Eichenwäldern fast vollständig, im
Osten werden sie dagegen zahlreicher, wohl aufgrund der ausgeprägteren
Sommerwärme.
Die
auffälligste floristische Trennlinie verläuft zwischen der
Subatlantischen und der Zentraleuropäischen Provinz. In der Atlantischen
Provinz sind a t l a n t i s c h e Arten wie Erica cinerea, Hyacinthoides non-scripta und Ceratocapnos claviculata
sowie atlantisch-subatlantische Arten zahlreich in den Wäldern
vertreten. In der Subatlantischen Provinz sind noch viele a t l a n t i s
c h - s u b - a t l a n t i s c h e Arten anzutreffen, beispielsweise Teucrium scorodonia, Hypericum pulchrum, Blechnum spicant, Luzula sylvatica, Galium saxatile, Lonicera periclymenum und Hedera helix.
Lianen scheinen – innerhalb der bodensauren Eichenwälder – in den atlantischen Territorien die größte Vitalität zu entfalten (Lonicera periclymenum, Hedera helix). In den südatlantischen Eichenwäldern treten zusätzlich die mediterran-atlantischen Arten Tamus communis und Rubia peregrina auf.
Der
weitaus überwiegende Teil der a t l a n t i s c h e n Arten (AT 8.1.,
8.1.S in Tab. 12) gehört zu den t h e r m i s c h o z e a n i s c h e n
Westseitenarten (vgl. JÄGER 1968). Es verwundert daher nicht, daß die
Bedeutung dieser Arten in den Kartierungseinheiten in Richtung ihres
klimatischen Optimums im Südwesten Eurasiens ständig zunimmt. Ulex europaeus,
Leitart des Arealtyps 8.1. (lusit-atl), ist eine typische thermisch
atlantische Art. Sie zeigt eine südatlantische Verbreitungstendenz, die
auch andere Arten mit ähnlichen Ansprüchen auszeichnet. Die Arten dieser
Gruppe erreichen häufig nicht das nordwesteuropäische Festland.
Auch
einige a t l a n t i s c h - s u b a t l a n t i s c h e Arten (AT
8.2., 8.5., 8(1).6.) erscheinen in den nordtemperat-euozeanischen
Bezirken Europas in bodensauren Eichenwäldern nur auf den Britischen
Inseln und fehlen auf dem Festland.
Einige
a t l a n t i s c h e Arten besiedeln nur die Südatlantische
Unterprovinz und teilweise mit Vorposten die Territorien der
Meridio-Atlantischen Provinzgruppe: Pyrus cordata, Pseudarrhenatherum longifolium, Daboecia cantabrica, Potentilla montana u.
a. Diese Arten werden unter dem hier neu etablierten Pseudarrhenatherum
longifolium-Arealtyp (8.1a. Nordlusit südatl, vgl. MEUSEL et al. 1965b,
K 44d) in der Tabelle zusammengefaßt. Die iberischen bodensauren
Eichenwälder innerhalb des 8.1a.-Areals erfahren noch eine Steigerung
ihres südatlantischen Charakters. Sie beherbergen neben weiteren
südatlantischen Arten zusätzlich eine Reihe von Endemiten (Omphalodes nitida, Crepis lampsanoides, Aquilegia vulgaris subsp. dichroa),
die ebenfalls dem AT 8.1a. zuzurechnen sind. Die größte Anzahl an
atlantischen Arten weist schließlich die nordwestiberische F14 auf.
Eine
andere Gruppe von Arten bewohnt ebenso Teile der Südatlantischen
Unterprovinz, aber in Iberien und südwärts davon nicht nur atlantische
Gebiete, sondern auch das weniger ozeanische Landesinnere und zeichnet
sich zusätzlich durch eine Präferenz der montanen Höhenstufe aus. Im
Gegensatz zum AT 8.1a. liegt der größte Teil des Areals dieser Arten in
der s u b m e r i d i o n a l e n Zone. Als Leitarten dieses
Verbreitungstyps können Quercus pyrenaica und Arenaria montana
gelten (JALAS & SUOMINEN (1976, 1983) Karte 305 und 688). Ihr Areal
entspricht dem westlichen Ausschnitt des westsubmediterranen Arealtyps
5.6. Ihr Arealtyp wird daher hier als Quercus pyrenaica-Arealtyp
5.6a. zusätzlich etabliert: (westmed)-westsubmed//mo-(südatl). Bei
ausschließlichen Gebirgspflanzen ist auch 6.1a. möglich. Wiederum
bewohnen einige Arten nur Segmente dieses 5.6a.-Areals, häufig handelt
es sich auch hierbei um iberische Endemiten.
Weitere s u b m e d i t e r r a n - o z e a n i s c h e Arten wie Luzula forsteri, Euphorbia amygdaloides, E. dulcis und Polystichum setiferum
sind gleichfalls auf die südatlantischen Einheiten konzentriert, können
aber in die Zentral- (F17, F21) und die Ostsubmediterrane Provinzgruppe
übergreifen. Ein häufiger Arealtyp ist 5.7.
M
e d i t e r r a n e Arten (ATG 1.) sind innerhalb der hier betrachteten
Kartierungseinheiten nur im Südwesten in den südatlantischen
Territorien vertreten, insbesondere in den planar-kollinen Einheiten.
Hierzu gehören beispielsweise Quercus
suber, Pinus pinaster, Erica arborea, Arbutus unedo, Tamus communis,
Rubia peregrina, Ruscus aculeatus, Asphodelus albus, Hypericum
androsaemum und Asplenium onopteris.
Den größten relativen Anteil dieser Arten enthält wiederum F14, es
folgen F7 und F15. Häufig ist der mediterran-atlantische Arealtyp 1.10.,
das heißt es handelt sich auch hier um ausgesprochen
thermisch ozeanische Arten.
Dagegen
finden sich in der Tabelle nur wenige Gefäßpflanzenarten, die im Norden
der Atlantischen Provinz in bodensauren Eichenwäldern ebenso gut oder
noch besser gedeihen als im Süden und in der Meridio-Atlantischen
Provinzgruppe. Hierzu kann man Galium saxatile, Blechnum spicant, Luzula sylvatica, Oreopteris limbosperma, Dryopteris aemula, eventuell auch Ilex aquifolium und Erica tetralix zählen. Viele dieser Arten sind im Osten der Subatlantischen Provinz Gebirgspflanzen.
Man kann daneben die meisten Vertreter der Mnium hornum-Moosgruppe als nordtemperat-atlantisch-subatlantisch betrachten.
Auf besonders armen Standorten sind ferner rohhumusbewohnende Moose wie Lophocolea heterophylla, Plagiothecium laetum und Aulacomnium androgynum
in den nordwestlichen Territorien der bodensauren Eichenwälder
bezeichnend. Die bodensauren Eichenwälder in Westirland und, in
geringerem Ausmaß, in Westbritannien sind besonders erwähnenswert. H y g
r i s c h h y p e r o z e a n i s c h e , austrocknungsempfindliche
Kryptogamen treten dort in einer Artenfülle auf, wie sie in bodensauren
Eichenwäldern sonst nicht wieder anzutreffen ist. Im Blechno-Quercetum petraeae
bei KELLY (1981) beträgt die mittlere Artenzahl (mAZ) 57,6.
Phanerogamen haben etwa 34 % Anteil an der mAZ, Kryptogamen dagegen 66
%. Die Einheit F1 repräsentiert, analog zur extrem thermisch
hyperozeanischen Einheit F14 in Iberien, eine extrem hygrisch
hyperozeanische Einheit innerhalb der hier behandelten Eichenwälder.
Charakteristisch sind Farne wie Hymenophyllum tunbrigense, H. wilsonii und Dryopteris aemula, weiter verbreitete ozeanische Bryophyten wie Lepidozia reptans und viele extrem atlantische Bryophyten wie Dicranum scottianum, aber insbesondere auch Lebermoose der Gruppe um Adelanthus decipiens. Diese Arten kommen auch in Lorbeerwäldern der Macaronesischen Unterregion (azor-canar-mad) vor.
Alle
für die atlantischen und subatlantischen Einheiten charakteristischen
ozeanischen Artengruppen fehlen den bodensauren Eichenwäldern in den
beiden östlichen Provinzen der Mitteleuropäischen Florenregion. Die
dortigen Kartierungseinheiten zeichnen sich vielmehr durch eine Zunahme b
o r e a l e r und e u r a s i s c h t e m p e r a t e r Sippen aus, die
bereits in den westlichen nordtemperaten Territorien einsetzt.
Vertreter dieser Artengruppe sind Pinus
sylvestris, Picea abies, Vaccinium myrtillus, Luzula pilosa,
Maianthemum bifolium, Vaccinium vitis-idaea, Trientalis europaea, Rubus
saxatilis, Pyrola rotundifolia, Calamagrostis arundinacea und Orthilia secunda.
Vielfach sind dies Nadelwaldelemente k o n t i n e n t a l e r
Arealtypen (AT 10.3., 10.8., 11.3.). Der osteuropäische Charakter wird
zusätzlich deutlich durch eine Reihe von Arten des sommergrünen
Breitlaubwaldes, die die Atlantische Provinz nicht erobert haben. Die
Arten des s u b k o n t i n e n t a l e n Asarum-Typs
8.11. mit Verbreitungsschwerpunkt im Osten der Mitteleuropäischen
Region meiden das Ulex-Areal 8.1.; auch die Arten mit den
subkontinentalen 8.13.- und 8.14.-Arealtypen sind hier anzuschließen
(vgl. Tab. 12, p. 5). Hierher gehören Acer platanoides, Euonymus verrucosa, Campanula persicifolia, Carex montana, Peucedanum oreoselinum, Potentilla alba
und zahlreiche weitere. Diese Artengruppen spielen auch in der im Osten
der Zentralsubmediterranen Provinzgruppe gelegenen illyrischen F21 eine
bedeutende Rolle, und die ebenfalls zentralsubmediterrane insubrische
F17 zeichnet sich durch das Nebeneinander von ozeanischen und
subkontinentalen Arten aus. Unter den Kryptogamen differenziert die
Artengruppe um Dicranum polysetum die osteuropäischen Einheiten.
Neben
den bereits erwähnten allgemein verbreiteten submediterranen Arten
zeichnen sich die zentraleuropäischen und sarmatischen
Kartierungseinheiten, insbesondere aber die südzentraleuropäische F20
und die illyrische F21 durch die Beteiligung charakteristischer
ostsubmediterraner Arten mit Arealerweiterungen ins östliche
Mitteleuropa aus: Genista tinctoria, Cytisus nigricans, Genista germanica und weitere. Doch können einige Vertreter des kontinentalen AT 5.9. Wie Vincetoxicum hirundinaria oder Tanacetum corymbosum weit nach Westen übergreifen, insbesondere in xerotherme Ausbildungen der F17 oder in die xerotherme subatlantische F18.
Die
besondere pflanzengeographische Bedeutung der s u b m e r i d i o n a l
e n Zone wird durch die Verbreitung der Eichenarten in den bodensauren
Eichenwäldern unterstrichen. In den nordtemperaten Territorien des
Areals sind Quercus robur und Q. petraea
die bestandsbildenden Arten. In den südtemperaten bis submediterranen
Territorien treten weitere Eichenarten in den bodensauren Eichenwäldern
auf, beispielsweise Q. pyrenaica, Q. canariensis und Q. suber auf der Iberischen Halbinsel oder Q. dalechampii, Q. polycarpa, Q. cerris und Q. frainetto in Südosteuropa.
Eine
Sonderstellung nimmt die illyrische F21 am südöstlichen Arealrand der
bodensauren Eichenmischwälder Europas ein. Sie ist durch zahlreiche e u -
s u b m e d i t e r r a n e Arten charakterisiert, die klimatisch
begünstigte Gebiete mit besonderer Eignung für anspruchsvolle
sommergrüne Breitlaubwälder in der Zentral- und Ostsubmediterraneis
bewohnen (häufig sind AT 5.4. und 5.5.): Quercus
cerris, Q. frainetto, Q. dalechampii, Q. polycarpa, Tilia tomentosa,
Acer tataricum, Fraxinus ornus, Hieracium racemosum, Potentilla
micrantha, Epimedium alpinum, Chamaecytisus hirsutus, Lathyrus venetus,
Silene viridiflora
und weitere. Viele dieser Arten treten in nur geringer Menge oder
Stetigkeit auf, doch im Überblick ist F21 durch das Neben- und
Miteinander der anspruchsvollen submediterran-nemoralen Elemente klar
gegen die südtemperate F20 differenziert.
Kartierungseinheiten
auf stark wechselfeuchten Standorten (F3, F9, F22) sind durch das
gehäufte Auftreten von nässetoleranten Arten gekennzeichnet,
beispielsweise durch Molinia caerulea, Deschampsia cespitosa, Agrostis stolonifera, Carex nigra und Lysimachia vulgaris. Für westliche Einheiten sind Myrica gale, Erica tetralix, Narthecium ossifragum und Sphagnum-Arten
bezeichnend, für zentraleuropäisch-sarmatische Molinia arundinacea und
Carex brizoides. Oreophyten differenzieren schließlich die montanen
Kartierungseinheiten. Hierbei spielen westsubmediterrane Gebirgspflanzen
besonders in F25 eine Rolle, während die zentralsubmediterrane F17
durch alpine Elemente ausgezeichnet ist (namentlich Luzula nivea und Phyteuma betonicifolium).
Stellung im pflanzensoziologischen System (Syntaxa) (J. Pallas)
Die Ordnung der azidophilen Eichenwälder, Quercetalia roboris Tüxen 1931, wird heute je nach
Auffassung in die Klasse Querco-Fagetea oder, wie ursprünglich, in eine eigenständige Klasse
Quercetea robori-petraeae Braun-Blanquet et Tüxen 1943 gestellt.
Die Ursache für die unterschiedliche Sichtweise liegt in der geographischen Ausgangssituation der Autoren, denn die Ordnung Quercetalia
enthält, wie auch Tabelle 12 zeigt, in den südtemperaten und
submeridional/montanen Territorien Europas wesentlich höhere Anteile an
anspruchsvolleren Laubwaldarten als beispielsweise im
nordwesteuropäischen temperaten Tiefland.
Die Quercetalia roboris
Tüxen 1931 beinhalten pflanzengeographisch heterogene Verbände, die
gleichsam nur durch die Vorherrschaft von Säurezeigern und durch die
Vorherrschaft azidotoleranter Eichen gekennzeichnet sind. Die
Kombination beider Merkmale ist das eigentliche Charakteristikum der
Ordnung.
Gegen
die atlantischen Einheiten, die in den extrem ozeanischen Gebieten
(Irland, Nordwestiberien) mit zahlreichen charakteristischen Arten
ausgestattet sind, nehmen sich bereits die subatlantischen Einheiten wie
Marginalsyntaxa aus und bleiben ohne eigene Kennarten. Die Grenze
zwischen Subatlantischer und Zentraleuropäischer Provinz tritt dagegen
wiederum durch den Ausfall der atlantisch-subatlantischen Arten recht
deutlich in Erscheinung. Doch wirken die zentraleuropäischen Einheiten
gegen die sarmatischen mit ihrem markanten Anteil an eurasischen oder
zirkumborealen Nadelwaldarten, eurasisch temperaten und osteuropäischen
Arten sowie ihren besseren Trophieverhältnissen wiederum nur wie
Marginalsyntaxa.
Im
Prinzip sind nur die Eichenwälder der beiden Mannigfaltigkeitszentren
(atl + submed) durch Azidophyten europäischer Verbreitung floristisch
positiv charakterisiert. Unter gesamteuropäischem Aspekt ergeben sich
daher synsystematische Probleme aus dem klimabedingten Rückgang der
atlantischen und atlantisch-subatlantischen Arten nach Osten. Zusätzlich
ist der größere Artenreichtum der submeridionalen Zone gegenüber der
temperaten zu berücksichtigen (vgl. MEUSEL & JÄGER 1989).
In Tabelle 12 (pp. 342-50 in BOHN et al. 2004) wird der Versuch unternommen, möglichst viele Einheiten der geographisch weitverbreiteten Ordnung Quercetalia roboris
miteinander zu vergleichen. Die folgende Übersicht stellt eine Revision
und Erweiterung des vom Verfasser (PALLAS 1996, 2000) bereits
publizierten Vorschlags dar. Die Einteilung basiert nunmehr
ausschließlich auf chorologischen Kriterien. Standortsunterschiede
(wechselfeuchte Einheiten) werden den geographischen Kriterien
untergeordnet.
Die
Sonderstellung der jeweils extrem ozeanischen Eichenwälder soll in
eigenständigen Verbänden deutlich werden. Diese Einheiten repräsentieren
jeweils Mannigfaltigkeitszentren. Das Hymenophyllo-Quercion umfaßt die hygrisch hyperozeanischen irischen und das Quercion robori-pyrenaicae
die thermisch hyperozeanischen Eichenwälder im Nordwesten Iberiens. Die
verbleibenden südtemperaten bodensauren Eichenwälder der Atlantischen
und Subatlantischen Provinz gehören zum Quercion roboris, und die verbleibenden nordtemperaten Eichenwälder der Atlantischen und Subatlantischen Provinz können im Molinio-Quercion
zusammengefaßt werden. In gleicher Weise werden auch die Eichenwälder
der Zentraleuropäischen und der Sarmatischen Provinz in einen
nordtemperaten Verband Vaccinio-Quercion petraeae und einen südtemperaten Verband Agrostio-Quercion zusammengefaßt.
Die zentral- und ostsubmediterranen bodensauren Eichenwälder gehören zum Castaneo-Quercion.
Quercetea robori-petraeae Braun-Blanquet et Tüxen 1943
Quercetalia roboris Tüxen 1931
1. Hymenophyllo-Quercion petraeae Pallas 2000: nordwesteuropäisch-temperat, mittelatlantisch, hygrisch hyperozeanisch
- F1 Quercetum petraeae Moss 1911 = Blechno spicant-Quercetum petraeae Br.-Bl. et Tx. 1952.
2. Quercion robori-pyrenaicae
(Braun-Blanquet et al. in P. Silva et al. 1950 corr. Br.-Bl. et al.
1956) Rivas-Martínez 1975: (submeridional)-südtemperat, südatlantisch:
nordlusit-galic-cant, thermisch hyperozeanisch. (Enthält iberische
Endemiten der AT 8.1a. und 5.6a., vgl. Tab. 12). Urprüngliche
Namensform: Quercion roboris broteroanae
Braun-Blanquet, Pinto da Silva, Rozeira & Fontes in Pinto da Silva,
Rozeira & Fontes 1950, Agronomia Lusitana 12(3): 435. Quercion occidentale Br.-Bl. et al. 1956 = Quercion roboris broteroanae (Braun-Blanquet et al. 1956, Agronomia Lusitana 18(3): 173).
- F14 Rusco aculeati-Quercetum roboris Braun-Blanquet, P. Silva et Rozeira 1956; Blechno spicant-Quercetum roboris Tüxen et Oberdorfer 1958
- F23 Myrtillo-Quercetum roboris Braun-Blanquet, P. Silva, Rozeira et Fontes in P. Silva, Rozeira et Fontes 1950 (Holotypus!)
- F24 Melampyro pratensis-Quercetum pyrenaicae Rivas-Martínez in Rivas-Martínez, T. E. Díaz, F. Prieto, Loidi et Penas
- 1984 (euskaldisch); Linario triornithophorae-Quercetum pyrenaicae Rivas-Martínez, T. E. Díaz, F. Prieto, Loidi et Penas 1984 (orokantabrisch)
- F25 Linario triornithophorae-Quercetum petraeae (Rivas-Martínez, Izco et Costa ex F. Navarro 1974) F. Prieto et Vázquez 1987; Luzulo henriquesii-Quercetum pyrenaicae (F. Prieto et Vázquez 1987) F. Prieto et Vázquez 1994.
Der Verband Quercion pyrenaicae Rivas Goday ex Rivas-Martínez 1964 [1963] umfaßt die bodensauren Quercus pyrenaica-Wälder mit Schwerpunkt in der Nordiberischen Provinz. Hierzu gehört in der Karte die Formationsuntergruppe G.4.1. mit den Einheiten G64 bis G70. Die Quercus canariensis-Wälder auf Silikatgestein der Formationsuntergruppe G.4.4. (G76, G77) stehen dieser Einheit ebenfalls nahe. Als submeridionale Einheit ist dieser Verband mit dem Castaneo-Quercion Soó 1964 zu vergleichen (siehe unten).
Wir
haben uns für eine Trennung der ehemaligen Unterverbände auf
Verbands-Rang entschieden. Die Unterschiede in der zonalen und der
Ozeanitätsbindung der beiden Verbände sollen deutlich werden: einerseits
temperat-südatlantische, meist von Quercus robur dominierte Wälder, andererseits submeridional-nordiberische, meist von Q. pyrenaica
dominierte Wälder (vgl. Karte). Dadurch wird vermieden, daß ein Verband
zwei Vegetationszonen umfaßt. In der Zukunft bleibt abzuklären, ob das
westsubmediterrane Quercion pyrenaicae und das zentral- bis ostsubmediterrane Castaneo-Quercion nicht in einer Ordnung submediterraner bodensaurer Eichenwälder vereinigt werden können.
3. Molinio caeruleae-Quercion roboris Scamoni et Passarge 1959: nordtemperat, mittelatlantische und nordsubatlantische Unterprovinz
- F2 W11 Quercus petraea-Betula pubescens-Oxalis acetosella woodland und W17 Quercus petraea-Betula pubescens-Dicranum majus woodland (excl. subcomm. a) in Rodwell 1991; Galio saxatilis-Quercetum Birse et Robertson 1976
- F3 W4 Betula pubescens-Molinia caerulea woodland, W16 Quercus-Betula-Deschampsia woodland, feuchte Ausbildungen, W17 Quercus petraea-Betula pubescens-Dicranum majus woodland, feuchte Ausbildungen, alle in Rodwell 1991.
- F9 Molinio caeruleae-Quercetum roboris (Tüxen 1937) Scamoni et Passarge 1959 (Lectotypus!)
- F8 Ilici-Quercetum roboris Tüxen 1930, Betulo-Quercetum roboris Tüxen 1930, Populo-Quercetum petraeae Tüxen 1951, Deschampsio flexuosae-Quercetum roboris Passarge 1966
- F10 Melico-Quercetum Björnstad 1971.
4. Quercion roboris Malcuit 1929: südtemperat, südatlantische und südsubatlantische Unterprovinz
- F26 Lathyro montani-Quercetum petraeae (Lapraz 1966) Rivas-Martínez 1983; Prunello hastifoliae-Quercetum petraeae Vigo ex Rivas-Martínez et Costa 1998
- F15 Hyperico pulchri-Quercetum roboris Rivas-Martínez, Báscones, Díaz, Fernández González et Loidi 1991.
- F7 Arbuto unedonis-Quercetum petraeae Lapraz 1963b; Lonicero periclymeni-Quercetum roboris Lapraz 1963b
- F5 Solidagini-Quercetum (Gaume 1924) Doing 1962 = Quercetum sessiliflorae Gaume 1924 nom. illegit. (Art. 31) (Holotypus!), syntax. Synon. Peucedano-Quercetum roboris Braun-Blanqet 1967. Der Verfasser verwirft an dieser Stelle seine frühere Gleichsetzung (PALLAS 1996: 21) des Teucrio scorodoniae-Quercetum petraeae Chouard 1925 mit dem Quercetum sessiliflorae Gaume 1924. Der Neotypus (Aufn. 19, Tab. 2, „Peucedano-Quercetum“, Rameau & Royer 1975, vgl. Pallas 1996: 21) wird beibehalten für das Quercetum sessiliflorae Gaume 1924, aber verworfen für das Teucrio-Quercetum Chouard 1925.
- F4 „Rusco-Quercetum“ Noirfalise ass. nov. prov. (nom. inval., Art. 3b)
- F6 Mespilo-Quercetum Frileux 1975
- F16 Teucrio scorodoniae-Quercetum petraeae Chouard 1925
- F18 Hieracio glaucini-Quercetum petraeae Lohmeyer 1978 corr. Denz 1994; Cladonio portentosae-Quercetum petraeae Pallas 1996.
- F8 Agrostio capillaris-Quercetum roboris Passarge 1968; Violo rivinianae-Quercetum roboris Oberdorfer 1957.
5. Vaccinio myrtilli-Quercion petraeae Pallas 1996: (nord)temperat (zentraleuropäisch, sarmatisch)
- F19, Calamagrostio-Quercetum (Hartmann 1934) Scamoni et Passarge 1959 (Holotypus!), Vaccinio vitis idaeae-Quercetum Oberdorfer 1957 p.p., typo excluso, zentraleuropäische Ausbildungen, „Pino-Quercetum“ sensu auct. German. non Reinhold 1944, non auct. polon.
- F12 Calamagrostio arundinaceae-Quercetum petraeae (Hartmann 1934) Scamoni et Passarge 1959, ostzentraleuropäische Ausbildungen. Die syntaxonomische Bewertung der osteuropäischen Einheiten ist wegen der großflächigen Durchmischung von Nadelwald und sommergrünem Laubwald nicht einfach. In diese natürlichen Übergänge greift noch zusätzlich der forstliche Einfluß des Menschen ein. Das zunächst für die Einheit F12 vorgesehene Syntaxon Querco-Pinetum J. M. Matuszkiewicz 1988 nom. inval. (Art. 5) et illegit. (Art. 31) ist jedoch in der Mehrzahl seiner Aufnahmen von Nadelhölzern dominiert und wurde daher folgerichtig vom Autor zum Dicrano-Pinion gestellt. Für die Etablierung der Syntaxa und die Wahl der nomenklatorischen Typen ist es jedoch von entscheidender Bedeutung, daß die Dominanz der Laubhölzer gewährleistet ist, denn sonst kann F12 nicht in der Laubwaldformation F verbleiben. Das Calamagrostio-Quercetum sollte als Leitgesellschaft der gesamten Zentraleuropäischen Florenprovinz betrachtet werden. Die Einheit F12 endet im Prinzip, wie auch das Areal von Quercus petraea, in der Zentraleuropäischen Provinz; sie erscheint daher in der Karte zu weit nach Osten ausgedehnt. Die sarmatische Einheit ist F13.
- F13 Serratulo tinctoriae-Quercetum roboris Pallas 2003 ass nov. hoc loco. Sarmatische, nadelholzhaltige bodensaure Stieleichenwälder. Holotypus, J. Pallas hoc loco: J. M. Matuszkiewicz 1988, Tab. 6, Aufn. 10 von T. Traczyk, aufgeführt unter „Serratulo-Pinetum“ nom. superfl. (Art. 29c).
Bei
der Einheit F13 stellt sich die gleiche Frage nach der Laubholzdominanz
zum Verbleib der Einheit in der Laubwaldformation. Die hier neu
beschriebene Assoziation repräsentiert den eichendominierten Teil der
fichtenhaltigen „subborealen“ Rasse des „Serratulo-Pinetum“.
6. Agrostio capillaris-Quercion petraeae Scamoni et Passarge 1959: südtemperat (südzentraleuropäisch, südsarmatisch)
- F11 Pyrolo-Quercetum petraeae Passarge 1957 (Lectotypus).
- F12 Pyrolo-Quercetum petraeae Passarge 1957, ostzentraleuropäische Ausbildungen.
- F13 Sarmatische Rasse des Serratulo-Pinetum (W. Matuszkiewicz et Polakowska 1955) J.M. Matuszkiewicz 1988 nom. superfl. (Art. 29c), als eichendominiertes Syntaxon noch neu zu beschreiben.
- F22 Molinio arundinaceae-Quercetum Samek 1962, Molinio arundinaceae-Quercetum roboris R. & Z. Neuhäusl 1967.
- F20 Luzulo luzuloidis-Quercetum petraeae Hilitzer 1932.
7. Castaneo-Quercion Soó 1964: submeridional (insb. zentralsubmediterrane Provinzgruppe)
- F17 Hieracio tenuiflori-Quercetum roboris Oberdorfer 1964
- F21 Castaneo-Quercetum (croaticum) Horvat 1938 (Lectotypus!), Erico-Quercetum petraeae Horvat 1959, Melampyro vulgati-Quercetum petraeae Puncer et Zupancic 1979 nom. illeg. [Art. 31], Luzulo forsteri-Quercetum petraeae Borhidi et Kevey 1996. Cytiso hirsuti-Quercetum petraeae (Stefanovic 1964) Pallas 2003 nom. nov. hoc loco pro nom. illegit. (Art. 34) Quercetum montanum illyricum Stefanovic 1964, Rad. Sumarsk. Fak. Sumarstvo Sarajevu 9(3), p. 22. Nomenklatorischer Typus (Lectotypus), J. Pallas hoc loco: op. cit., Tab. II, p. 26-29, Aufn. 8. Nicht gesondert herausgestellt werden die von verschiedenen Autoren unter „Genisto pilosae-Quercetum“ beschriebenen besonders xerophytischen Felspflanzen-Eichenwälder. Sie kommen im Areal von F20, F21 und auch F17 vor und vermitteln, insbesondere auf etwas basenreicheren Felsstandorten, zur Formation G. Sehr ähnlich ist Einheit F18 in der Subatlantischen Provinz.
Gliederung in Untereinheiten (J. Pallas)
In der Gesamtlegende ist die Hauptgliederung der azidophilen Eichenwälder nach Höhenstufen
erfolgt, die im wesentlichen von Norden nach Süden angeordnet sind:
- planar-kolline Ausbildungen (F1-F13), (auf der Iberischen Halbinsel nicht vorhanden - nur östlich der Pyrenäen)
- kollin-submontane Ausbildungen (F14-F22),
- montan-hochmontane Ausbildungen (F23-F26).
Subformation F1
| |
Untergruppe F1.2 (kollin-submontan)
| 
Untergruppe F1.3 (montan-hochmontan)
|
Iberische Kartierungseinheiten von Untergruppe F.12

Iberische Kartierungseinheiten von Untergruppe F1.3 von F1
Subformation F2
Charakterisierung und typologische Abgrenzung; geographische Verbreitung
Nach BOHN et al. (2002) liegt das Verbreitungsgebiet dieser ozeanischen Formation mit den Hauptbaumarten Fraxinus excelsior, Quercus robur, Ulmus glabra und Quercus petraea
liegt überwiegend auf den Britischen Inseln, ferner am Fuß und im
Innern der westlichen Pyrenäen und des Kantabrischen Gebirges; hinzu
kommen isolierte Vorkommen entlang der Westküste von Norwegen und in
Nordwestfrankreich.
Die
Bestände der Formation besiedeln in der Regel basenreiche, oft
kalkhaltige, mäßig saure bis neutrale Braunerden. Charakteristisch ist
eine relativ artenreiche meso- bis eutraphente Flora im Unterschied zur
vorhergehenden Formation der artenarmen, azidophilen Eichen- und
Eichenmischwälder (F.1). Eichen-Eschenmischwälder kommen hauptsächlich
außerhalb des natürlichen Areals von Eichen-Hainbuchenwäldern und
Buchenwäldern vor, wobei Fagus sylvatica
in Spanien, Frankreich und Südengland allerdings örtlich als
natürliches Element in dieser Formation vertreten ist. In Irland nimmt Fraxinus excelsior
alle basenreichen, mäßig trockenen bis feuchten Standorte als
herrschende Baumart ein, da hier Buche und Hainbuche von Natur aus
fehlen.
 | |

F33 - Cantabrian-Euscaldian mixed oak-ash forests (Fraxinus excelsior, Quercus robur, Quercus petraea, Tilia platyphyllos, Fagus sylvatica) with Polystichum setiferum, Pulmonaria affinis, Stellaria holostea, Potentilla sterilis
|
Subformation F5
Natürliche autochthone Vorkommen von Buchenwald (Fagus sylvatica) gibt es in der Serra da Estrela nicht. Auch eiszeitliche Vorkommen der Rotbuche konnten durch Pollenanalysen für die Serra da Estrela nicht belegt werden, obwohl Simulationen unter den Klimabedingungen von vor ca 20.000 Jahren ein prähistorisches Vorkommen zu dieser Zeit in der Serra da Estrela möglich erscheinen lassen.
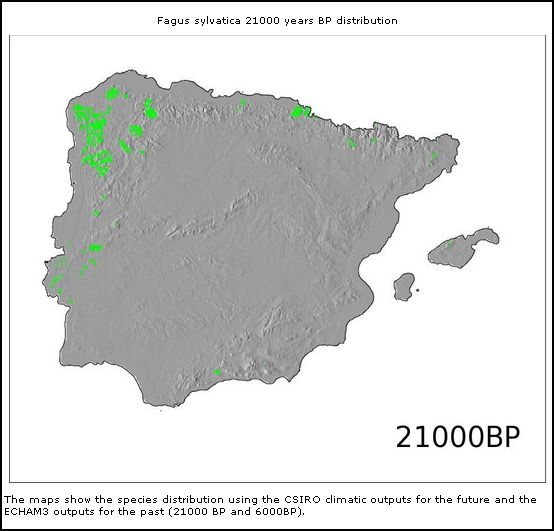
Für den Norden der Iberischen Halbinsel ist die Rotbuche als jedoch als autochthone Spezies nachgewiesen und ihre rezente Verbreitung im Norden der Iberischen Halbinsel ist auch aus prähistorischen Zeiten durch Pollenanalysen belegt. In der Karte der Potentiellen Natürlichen Vegetation Europas erscheint die Buche in Form verschiedener Buchenwaldgesellschaften in der Subformation F5 (Kartierungseinheiten (F98, F99) und (F136, F137, F138)) auf der Iberischen Halbinsel (BOHN et al. 2002, pp. 286).
F.5 Buchenwälder und Buchenmischwälder
Charakterisierung und typologische Abgrenzung
Nach BOHN et al. (2002) sind Buchenwälder durch die natürliche Vorherrschaft der Buche (Fagus sylvatica)
in der Baumschicht gekennzeichnet; deren Deckung beträgt hier
mindestens 50 %, meist jedoch über 90 %. Buchenwälder weisen nur wenige
Mischbaumarten in der 1. oder 2. Baumschicht auf, da die Buche in ihrem
klimatischen und edaphischen Optimalbereich – auf „Normalstandorten“ –
außerordentlich konkurrenzkräftig und gegenüber anderen Baumarten
unduldsam ist; dabei kommen ihr die hohe Schattenverträglichkeit und
weite Standortsamplitude zugute, die von nährstoffarmen bis -reichen
Böden, von betont frischen oder staufeuchten bis zu trockenwarmen
Standorten und vom Tiefland bis in die hochmontane Stufe der Gebirge
reicht. Mischbaumarten erlangen gewöhnlich erst in standörtlichen
Grenzbereichen der Buchenwälder, im Übergang zu anderen
Waldgesellschaften extremerer Standorte, höheren Anteil. Die Beteiligung
von Mischbaumarten hängt von der Trophie der Standorte, der
Bodenfeuchte, der Höhenlage und von der geographischen Lage sowie der
Gebietsflora ab. Die wichtigsten Mischbaumarten sind Quercus petraea (bodensaure und staufeuchte Tieflagenstandorte), Carpinus betulus (reichere Tieflagenstandorte, 2. Baumschicht), Fraxinus excelsior, Acer pseudoplatanus, A. platanoides, Ulmus glabra (reichere sowie blockreiche Standorte vor allem höherer Lagen), Abies alba, Picea abies
(vorwiegend in höheren Mittel- und Hochgebirgen). Die Waldbestände sind
in der Regel zwei-, seltener dreischichtig, wobei Strauch- und
Moosschicht weitgehend fehlen. Zusammensetzung und Deckung der Kraut-
(und Strauchschicht) variieren sehr stark in Abhängigkeit von der
Nährstoff- und Wasserversorgung des Bodens, ferner von Exposition,
Meereshöhe und geographischer Lage: Danach werden u. a. azidophile,
basiphile und thermophile, artenarme und -reiche, moosreiche,
simsenreiche, seggenreiche, grasreiche, krautreiche, farnreiche und
hochstaudenreiche Ausbildungen unterschieden, die verschiedenen
Verbänden innerhalb der Querco-Fagetea bzw. Fagetalia sylvaticae angehören.
Subformation F5 auf der Iberischen Halbinsel
| |

Iberischer Anteil der Untergruppe F5.1
| 
Iberischer Anteil der Untergruppe F5.2
|
Die wichtigsten Kartierungseinheiten der Formation F nord-nordwestlich und nördlich von der Serra da Estrela:

F14 - Galician-north Lusitanian hyperoceanic pedunculate oak forests (Quercus robur, partly Quercus pyrenaica, Quercus suber) with Laurus nobilis, Viburnum tinus, Pyrus cordata, Daboecia cantabrica, Andryala integrifolia
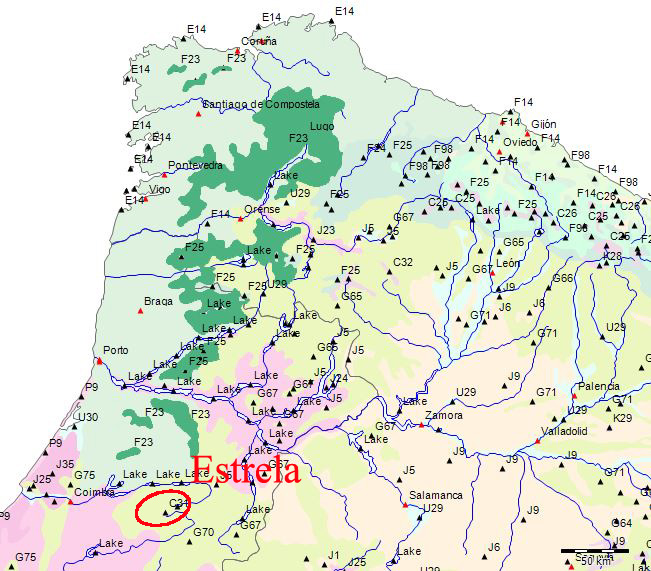
F23 - Galician-north Lusitanian oak forests (Quercus robur, Quercus pyrenaica) with Betula pubescens subsp. celtiberica, Cytisus striatus, Dryopteris aemula, Anemone trifolia subsp. albida, Omphalodes nitida
Formation J
An den südöstlich und nordöstlichen Rändern der Serra da Estrela finden wir die Formation J der Mediterranen Hartlaubwälder und -gebüsche.
J - Mediterrane Hartlaubwälder und -gebüsche[87]
Charakterisierung und typologische Abgrenzung; Bestandesstruktur und Physiognomie
Die Formation „Mediterrane Hartlaubwälder und -gebüsche“ umfaßt Gesellschaften xeromorpher immergrüner Baum- und Straucharten, vor allem der Gattungen Quercus (Subgenus Sclerophyllodrys), Pinus, Juniperus, Olea und Pistacia.
Dominanzen einer Baum- oder Strauchart sind häufig. Hartlaubwälder und
-gebüsche kommen auf ganz verschiedenen Substraten und in allen
Expositionen vor, doch werden grundwasserbeeinflußte Standorte gemieden.
Die
Struktur der Bestände hängt von abiotischen Standortfaktoren und der
Artenzusammensetzung ab, vor allem aber von der aktuellen und früheren
Nutzung. Die meisten immergrünen Laubwaldarten können – je nach den
äußeren Einflüssen, manche anscheinend auch genetisch prädisponiert –
baum- oder strauchförmig wachsen. Bestände mit dichtem Kronenschluß sind
licht- und artenarm. Der spärliche Unterwuchs solch dichter Wälder und
Gebüsche besteht aus vereinzelten Kleinsträuchern, Gräsern, Lianen,
Geophyten, Farnen sowie dem Jungwuchs der Bäume und Sträucher. Viel
häufiger sind lichte Bestände, wie sie von Natur aus in Steillagen, in
der Regel aber als Folge von Nutzungseinflüssen wie Holzentnahme und
Beweidung vorkommen. Durch ungeregelte Holznutzung degradierte Bestände
haben Gebüsch- oder Buschwaldstruktur (Macchia, maquis). Sind sie, meist
durch Beweidung, fleckweise aufgelöst und mosaikförmig mit
Zwergstrauch- und krautiger Vegetation verzahnt, so sprechen wir von
Garrigue. Kernwüchsige Weidewälder haben meist geringen Kronenschluß,
unausgewogene Altersstruktur und eine durch Ökotoneffekte reiche
Binnenstruktur.
Mediterrane
Hartlaubwälder sind in für den Ackerbau günstigen Räumen schon während
des Neolithikums, in ungünstigeren Lagen vor allem in Zeiten hohen
Bevölkerungsdrucks durch extensive ungeregelte Holzentnahme und
Beweidung mit Ziegen und Schafen zerstört oder degradiert worden. An
ihre Stelle traten verschiedene Kulturlandformationen. Bei extensiver
Beweidung entstanden sekundäre Kleinstrauchgesellschaften (regional
unterschiedlich als Tomillares, Phrygana oder Batha bezeichnet), bei
anhaltender regressiver Syndynamik geophyten- und annuellenreiche
Trockenrasen und Heiden.
In
jüngerer Zeit beobachtet man in den meisten Ländern am Nordrand des
Mittelmeeres bei nachlassendem Nutzungsdruck sich selbst überlassene
regenerative Hartlaubvegetation. Das dynamische Potential solcher
Bestände, besonders der zeitliche Ablauf der progressiven Sukzession und
die jeweilige Schlußwaldgesellschaft, sind in hohem Maße
standortabhängig und im einzelnen wenig bekannt.
Geographische Verbreitung
Das natürliche Wuchsgebiet mediterraner Hartlaubwälder und -gebüsche in Europa umfaßt den
größten
Teil der Iberischen Halbinsel, die Tieflagen und küstennahen Gebiete
Südfrankreichs, Italiens, der Adria-Länder und der südlichen
Balkanhalbinsel sowie die Mittelmeerinseln von den Balearen bis zur
Ägäis mit Ausnahme einiger Gebirgs- und Küstenräume (s. Karte 14 im
Textband sowie die Blätter 7 und 8 der Europakarte). Während mediterrane
Hartlaubwälder im iberischen Teil des Verbreitungsgebietes auch weit im
Inland vorkommen, bleiben sie im Norden des Areals auf einen meist
wenige Kilometer schmalen Küstensaum beschränkt, können aber im Süden
(Sizilien, Mittelgriechenland, Peloponnes) bis zu 70 km landeinwärts
reichen. Das Areal immergrüner mediterraner Hartlaubvegetation setzt
sich außerhalb Europas nach Süden (Teile der Nordküsten Marokkos,
Algeriens, Tunesiens, Libyens) und Osten fort (Küstenräume der West- und
Südtürkei, Zyperns, Syriens, des Libanon und Israels).
Floristische Zusammensetzung (Artengefüge)
Die Baumartenzusammensetzung mediterraner Hartlaubwälder ist meist ziemlich einförmig. Nur
eine Art dominiert in der Regel den Kronenraum, oft eine der immergrünen Eichenarten (Subgenus Sclerophyllodrys). Die Stein-Eiche (Quercus ilex)
konkurriert am erfolgreichsten in subhumiden Gebieten; sie ist auf der
Iberischen Halbinsel außerhalb Kantabriens und Kataloniens vertreten
durch Q. ilex subsp. rotundifolia (= Q. rotundifolia, Q. ilex subsp. ballota), im übrigen Mittelmeergebiet durch Q. ilex subsp. ilex. In trockeneren und etwas winterkälteren Lagen wird Q. ilex durch die Kermes-Eiche (Quercus coccifera) ersetzt. Diese ist gegenüber Verbiß toleranter als die Stein-Eiche. Quercus coccifera ist Waldbildner besonders im Ostmediterrangebiet (und wird hier von manchen Autoren traditionell unter dem Synonym Q. calliprinos
nomenklatorisch abgegrenzt), während sie im Westen kaum höher als 2 m
wird. In mesomediterranen Gebieten und vor allem im Übergang zu
submediterranen Klimaräumen können auch laubabwerfende Baumarten stärker
beteiligt sein: im Westen besonders Eichenarten wie Quercus faginea und Q. pubescens, im Osten Q. cerris, Q. frainetto, Fraxinus ornus und Carpinus orientalis. Eichen-Weidewälder und Macchien mit anthropogen geringem Eichenanteil haben oft eine dichte immergrüne Strauchschicht, die aus Erica arborea, Arbutus unedo oder A. andrachne gebildet sein kann. An humiden Standorten finden sich Laurus nobilis und Myrtus communis. Viel weniger verbreitet als die Gesellschaften mit Quercus ilex oder Q. coccifera sind Wälder aus Quercus-Arten der Untergattung Cerris. Gleichwohl sind sie gebietsweise landschaftsbestimmend, etwa die Kork-Eiche (Quercus suber) in Südwestspanien
und Portugal und Q. ithaburensis subsp. macrolepis in Südostitalien und Süd- und Westgriechenland.
In
thermomediterranen Gebieten sind die immergrünen Eichen auf
Sonderstandorte oder klimatisch gemäßigte Berglagen beschränkt; hier ist
auf sandigen Böden in Küstenlagen und auf Kalkgestein oft Pinus halepensis die vorherrschende Baumart (in der Ägäis und im mediterranen Kleinasien P. brutia). Durch ihre widerstandsfähige Borke und das hohe Regenerationspotential durch Samenkeimung tolerieren P. halepensis und P. brutia
Feuereinwirkung mehr als andere Baumarten und profitieren so von
Waldbränden. Bei häufigen Bränden vermögen die Kiefernarten auch in
mesomediterranen Gebieten die Eichen als Hauptbaumarten zu verdrängen.
Die in vielen Gebieten häufigste Strauchart der thermomediterranen Hartlaubgebüsche ist – mit oder ohne Pinus-Schirm – Pistacia lentiscus, meist als Strauch von unter 2 m Höhe, selten als Baum von bis zu 6 m. Sie wächst oft zusammen mit der Wildform des Ölbaums (Olea europaea subsp. oleaster) und dem Johannisbrotbaum (Ceratonia siliqua).
Olea wie Ceratonia gehören nach heutigem Kenntnisstand nicht zur
ursprünglichen europäischen Flora, sondern wurden in ihren nahöstlichen
Indigenaträumen in Kultur genommen und von dort aus im Mittelmeergebiet
verbreitet, wo sie in der Folge verwilderten und seit langem in der
natürlichen Vegetation fest eingebürgert sind. Weitere wichtige
bestandsbildende und weit verbreitete Holzarten sind im
Thermomediterranraum Juniperus phoenicea (incl. subsp. turbinata), meist küstennahe Gebüsche bildend, und J. oxycedrus subsp. macrocarpa, oft baumförmig Küstensande besiedelnd.
Während
anthropogen aufgelichtete Hartlaubwälder auf kleiner Fläche eine hohe
Artenzahl aufweisen können, sind wenig oder lange nicht mehr gestörte
Bestände dicht und auffällig artenarm. Besonders schattentolerant und
daher unter dem Schirm dichter Hartlaubwälder weit verbreitet sind
Farnartige (Asplenium onopteris, Selaginella denticulata), immergrüne, teils klimmende Halbsträucher und Lianen (Ruscus aculeatus, Asparagus acutifolius, A. aphyllus, Smilax aspera, Rubia peregrina, R. tenuifolia) sowie Geophyten (Cyclamen-Arten, Arisarum vulgare und andere Araceae). Im Schutz niedriger Hartlaubgebüsche wachsen dagegen Horstgräser wie Piptatherum miliaceum, P. coerulescens und Ampelodesmos mauritanica, deren Halme die Sträucher überragen können.
Stellung im pflanzensoziologischen System (Syntaxa)
Die Formation „Mediterrane Hartlaubwälder und -gebüsche“ fällt im wesentlichen mit der pflanzensoziologischen Klasse Quercetea ilicis zusammen. (In diese Klasse gehören obendrein bestimmte oromediterrane Nadelwälder wie die Cupressus-Wälder Kretas, die in der Europakarte zur Formation K gezählt werden.) Mesomediterrane Quercus- und Pinus-Wälder werden zur Ordnung Quercetalia ilicis zusammengefaßt. Die Ordnung Pistacio lentisci-Rhamnetalia alaterni umfaßt Gebüschgesellschaften sowie Pinus- und Juniperus-Bestände
vorwiegend im thermomediterranen Bereich. Die weitere Gliederung in
Verbände und Assoziationen ist in Teilgebieten (z. B. Spanien)
vorgenommen worden, jedoch fußen die meisten Vorschläge auf relativ
geringer Datenbasis, und keiner berücksichtigt alle europäischen
Teilgebiete. Der Verband Oleo-Ceratonion
gilt seit langem als bezeichnende Einheit der aktuellen
höchstentwickelten thermomediterranen Vegetation. Einem jüngeren
Vorschlag zufolge soll dieser Verband auf das westliche Mittelmeergebiet
beschränkt bleiben und gegen ein östliches Pendant, Ceratonio-Rhamnion,
abgrenzbar sein. Für eine anzustrebende, zirkum-mediterran stimmige
Synsystematik der südeuropäisch-nordafrikanisch-nahöstlichen
Hartlaubvegetation bedarf es einer gründlichen Revision und Emendierung
der Mehrzahl der vorhandenen syntaxonomischen Regionalkonzepte.
Makroklimatische Gegebenheiten
Hartlaubwälder
und -gebüsche wachsen in Gebieten mit ausgeprägtem mediterranem
Winterregenklima, also bei trockenen warmen Sommern und kühlen feuchten
Wintern. Strenge Fröste fehlen, in thermomediterranen Gebieten sind
Fröste generell selten. Die Niederschlagsmaxima liegen gewöhnlich im
November/Dezember und im Februar/März. Die Sommermonate – in den
trockenen südlichen Gebieten schon ab Mai und bis einschließlich
September – bringen wenig oder keinen Niederschlag. Bei ausgeprägter
Reliefgestaltung sorgen Luv-Lee-Effekte für erhebliche lokalklimatische
Differenzierung. Hinzu kommen Schwankungen in der Verteilung und den
Jahressummen der Niederschläge. Die mittleren jährlichen
Niederschlagssummen in Meereshöhe liegen zwischen 400 und 900 mm,
seltener über 1200 mm (z. B. Kerkira) oder unter 400 mm
(südostspanisches Trockengebiet, Südostkreta). Die Temperaturmittel des
wärmsten Monats liegen zwischen 25 und 28 °C, die des kältesten Monats
zwischen 6 und 13 °C. Besonders auf den kleineren Inseln und an
exponierten Küsten ist der Windfaktor vegetations- und strukturprägend:
Er mindert die Wettbewerbskraft hygrisch anspruchsvoller immergrüner
Holzarten und verhindert örtlich – auch ohne anthropogene Einflüsse –
baumförmigen Wuchs, so daß die Konkurrenzfähigkeit niedriger Gebüsche
und von
Buschwäldern gefördert wird.
Rolle im Landschaftsgefüge, Erhaltungszustand, Landnutzung; Naturschutz
Hartlaubwälder
gelten oft als die zonale Vegetation des Mittelmeergebiets schlechthin,
doch deuten pollenanalytische Befunde auf eine Zunahme immergrüner
Eichenwälder erst im Gefolge der neolithischen Landnahme hin. Generell
ist im zirkum-mediterranen küstennahen Raum von einer
prähistorisch-historischen, anthropogen-edaphischen Verschiebung des
Konkurrenzgleichgewichtes zwischen immergrünen und winterkahlen
Schlußgesellschaften zugunsten der immergrünen Hartlaubvegetation
auszugehen. Freilich sind die Untersuchungen zu lückenhaft, um eine
differenzierte Einschätzung der Vegetationsgeschichte für alle Teilräume
vornehmen zu können. Auch darf die heute standortgemäße (potentielle)
natürliche Vegetation nicht mit der vom Menschen noch wenig beeinflußten
des Neolithikums gleichgesetzt werden. Erhebliche Bodenerosion – mit
und ohne Zutun des Menschen – hat die standörtlichen Voraussetzungen für
Waldwuchs und -struktur gründlich verändert. Eine standortgemäße
Regeneration von Wäldern in Bereichen, wo diese degradiert oder
verschwunden sind, hängt von Bodenbeschaffenheit und Wasserversorgung,
vom Relief und lokalen Klima, dem Diasporenangebot und von eventuellen
Störungen ab. Beobachtungen an fortgeschrittenen Sukzessionsbeständen
von Hartlaubwäldern in Südfrankreich lassen auf ihre Weiterentwicklung
in
Richtung einer stärkeren Beteiligung laubabwerfender Eichen (hier
Quercus pubescens) schließen. Auch diese bestandesdynamischen Befunde
sind aber bisher nur regional abgesichert und dürfen nicht unkritisch
auf das gesamte Hartlaubwald-Gebiet verallgemeinert werden. Das
Regenerationspotential von Hartlaubgehölzen ist in mesomediterranen
Gebieten im allgemeinen besser als in den thermomediterranen Regionen.
Der Erhaltungszustand mediterraner Hartlaubwälder und -gebüsche ist für die einzelnen zonalen
Typen unterschiedlich, für die meisten Waldgesellschaften aber – trotz großräumig progressiver
Sukzessionstrends
– schlecht. Manche Gesellschaften sind als einigermaßen repräsentativer
ungestörter Wald praktisch unbekannt, die Schlußgesellschaft der
betreffenden Kartierungseinheit infolgedessen hypothetisch. Für manche
Standorte, besonders solche, die sich für den Anbau von Kulturpflanzen
eignen, sind keinerlei Waldreste belegt. Die meisten Bestände von alten
Hartlaubwäldern sind strukturell mehr oder minder stark überformt und
auf bestimmte, für den Menschen schwer erreichbare oder sonst ungünstige
Standorte beschränkt. Dies erschwert den Blick auf die potentielle
standörtliche Variabilität der Gesellschaften. Langfristige Forschung
auf der Grundlage systematisch angelegter Dauerflächen zur Erhellung der
Sukzessionsvorgänge fehlt weithin bzw. scheitert gewöhnlich an der
Zerstörung der experimentellen Einrichtungen. Pinus-dominierte Wälder
tragen zudem ein hohes Feuerrisiko in sich. Ebene Standorte mit gut
entwickelten und tiefgründigen Böden stehen für das Studium der zonalen
Waldentwicklung in der jahrtausendealten Kulturlandschaft des
Mittelmeerraumes kaum zur Verfügung. Gering beeinflußte reife
Hartlaubwälder sind wegen ihrer Seltenheit für Naturschutz und
Wissenschaft von großer Bedeutung. Dasselbe gilt wegen ihres
Entwicklungspotentials auch für aus Niederwaldnutzung hervorgegangene
fortgeschrittene Regenerationsbestände und für ausgedehnte kernwüchsige
Weidewälder. Aus Naturschutzsicht
sind
freilich Hartlaubwälder nicht generell ihren Ersatzgesellschaften
vorzuziehen, welche ungleich reicher an Organismen und Kleinhabitaten
sind als die zonale Schlußgesellschaft.
Gliederung in Untereinheiten
Die
Hartlaubwälder und -gebüsche des Mittelmeerraumes lassen sich zunächst
nach ihrer klimatischen Präferenz in eine Gesellschaftsgruppe
mesomediterraner und eine Gruppe thermomediterraner Verbreitung
differenzieren. Innerhalb jeder dieser beiden Gesellschaftsgruppen ist
die vorherrschende Baumart – meist aus der Gattung Quercus – vorrangiges
Kriterium der weiteren Differenzierung.
Diese
bestimmt als Matrixart in erheblichem Maß den Bestandescharakter. Die
weitere Unterteilung in Kartierungseinheiten fußt dagegen hauptsächlich
auf substratbedingten und chorologischen Spezifika des Unterwuchses.
Ein
rascher Überblick über die standörtlich differenzierte regionale
Verteilung der insgesamt 53 Kartierungseinheiten läßt sich am besten
anhand ihrer drei großen südeuropäischen Teilareale – der Iberischen
Halbinsel mit den Balearen und Südfrankreich, der Apenninhalbinsel mit
Sizilien, Sardinien und Korsika und der Balkanhalbinsel mit Kreta und
den ägäischen Inseln – gewinnen (vgl. Karte 14). Dabei wird aus Gründen
der Übersichtlichkeit und Straffung der Darstellung vornehmlich auf die
natürlichen (in vielen Fällen hypothetischen und aus standörtlichen und
syndynamischen Befunden abgeleiteten) Schlußgesellschaften eingegangen,
dagegen nicht oder nur punktuell auf die aktuell landschaftsprägende
Ersatzvegetation. Zur besseren Orientierung ist die Legendennummer der
jeweiligen Kartierungseinheit in Klammern beigefügt.
Iberische Halbinsel und Balearen, Südfrankreich
Am
großflächigsten und am stärksten differenziert zeigen sich die
Hartlaubwälder und -gebüsche in Spanien und Portugal. Nur der
atlantische Norden und Nordwesten der Iberischen Halbinsel wird nicht
von ihnen besiedelt, mit Ausnahme einiger reliktischer Waldinseln mit Quercus ilex subsp. ilex (J13),
die in K a n t a b r i e n bei Bilbao und Santander in thermisch
begünstigten Tälern mit Eichen-Eschenwäldern verzahnt sind, sowie
lokaler Quercus ilex subsp. rotundifolia-Karstwälder (J11),
die auf edaphischen Sonderstandorten in die Buchen- und
Eichenmischwald-Landschaft der mittleren kantabrischen Küstenkordillere
kleinräumig eingestreut sind.
Mi t t e l - u n d Sü d p o r t u g a l ist das Hauptverbreitungsgebiet der südwestiberischen Korkeichenwälder (Quercus suber).
Ihr thermomediterraner Flügel besiedelt zum einen küstennahe,
nährstoffarme Sandböden des Alentejo und der Küstenlandschaft
Mittelportugals (J35), hier u. A. differenziert durch den psammophilen Juniperus navicularis, zum anderen skelettreiche Böden über festen Silikatgesteinen (J36)
ohne ausgesprochene Psammophyten. Mit zunehmender Küstenferne und
Höhenlage (Serra de Monchique, Portugiesisches Scheidegebirge) stellen
sich mesomediterrane Ausbildungen der Korkeichenwälder ein, die keine
thermisch anspruchsvollen Holzarten (Olea, Myrtus) mehr enthalten; dafür können hier laubabwerfende Eichen (Quercus canariensis) beigemischt sein (J25).
Die geschlossene portugiesische Korkeichen-Landschaft erstreckt sich
von der Algarve im Süden bis in die Gegend von Coimbra; nördlich davon
besteht noch ein isoliertes mesomediterranes Teilareal im mittleren
Duero-Becken im Hinterland von Porto mit Juniperus oxycedrus als
Begleit-Holzart (J24). Die nördlichsten Korkeichenwälder mit Reliktcharakter (J23)
– differenziert durch Arten mesophiler Laubwälder in der Krautschicht –
finden sich im spanischen Galicien im Tal des Sil bei Orense. Mit
abnehmender Ozeanität des Klimacharakters, so im Einzugsbereich der
Guadiana (Alto Alentejo), ziehen sich die Korkeichenwälder ostwärts
inselartig auf hygrisch günstige Standorte zurück und werden von
mesomediterranen Quercus ilex subsp. rotundifolia-
Wäldern
abgelöst. In den wenigen Kalksteingebieten des thermomediterranen
Portugal wird die Korkeichen-Landschaft durch endemitenreiche, basiphile
Kermeseichen-Wacholder gebüsche (Quercus coccifera, Juniperus phoenicea subsp. turbinata) unterbrochen, so bei Lissabon und in der Serra de Arrabida bei Setúbal (J29) sowie auf Jurakalken an der Südwestspitze der Iberischen Halbinsel am Cabo de São Vicente (J41). Die Quercus ilex subsp. rotundifolia-Wälder in den Kalkgebieten des Hügellandes der südlichen Algarve (J38) bilden die westlichen Ausläufer der andalusischen Steineichenwälder.
Die thermomediterranen Quercus suber-Wälder des lusitanischen Typs (J35, J36)
erstrecken sich in den Silikatgebieten im küstennahen S ü d s p a n i e
n über Gibraltar hinaus bis in die Gegend von Marbella. Ihr
mesomediterraner Flügel im Bergland bei Algeciras oberhalb ca. 400 m (J26) ist jedoch von portugiesischen mesomediterranen Ausbildungsformen deutlich floristisch unterschieden (z. B. Teucrium pseudo-scorodonia, Luzula forsteri subsp. baetica).
Die thermomediterranen Kalkgebiete des Guadalquivir-Beckens und entlang
der Küste von Malaga bis Motril sind dagegen das Wuchsgebiet
südwestiberischer Quercus ilex subsp. rotundifolia-Wälder (J38), ebenfalls mit eigenen floristischen Zügen (u. a. Aristolochia baetica, Bupleurum gibraltarium). Quercus ilex subsp. rotundifolia
ist im thermomediterranen Andalusien durchaus auch Waldbildner auf
silikatischen Substraten, so nördlich des Guadalquivir in der Sierra
Morena (J37), wo Quercus suber
aus klimatischen Gründen nicht mehr wettbewerbsfähig ist. Einen
edaphischen Sonderfall stellt das ausgedehnte andalusische
Schwarzerdegebiet bei Cadiz dar, in welchem auf Grund des starken
Anteils schwellfähiger Tonmineralien im Boden Quercus-Arten nicht konkurrieren können und deshalb Olea europaea subsp. oleaster als Hauptwaldbildner vermutet wird (J42). Die mesomediterrane Stufe Andalusiens östlich des Guadalquivir-Beckens (ca. 800-1500 m) ist die Domäne basiphiler Quercus ilex subsp. rotundifolia-Wälder, auf Böden mit größerer Feuchtigkeit mit Quercus faginea und Acer monspessulanum, die u. a. durch das Vorkommen von Paeonia coriacea von thermomediterranen Ausbildungsformen differenziert sind (J3). Das Hochgebirge der Sierra Nevada beherbergt gar noch supramediterrane Quercus ilex subsp. rotundifolia-Wälder bei 1400-1900 m, floristisch differenziert durch Begleit-Holzarten wie Acer granatense, Berberis vulgaris subsp. australis und Prunus ramburii (J10).
Das
s ü d o s t s p a n i s c h e T r o c k e n g e b i e t zwischen
Almeria und Murcia, im Regenschatten der bätischen Hochgebirge gelegen,
ist bei einem Jahresniederschlag von unter 400 mm das potentielle
Wuchsgebiet semiarider Pistacia lentiscus-Buschwälder mit Olea europaea subsp. oleaster und örtlich Juniperus phoenicea als Begleit-Holzarten sowie wenigen Lianen bzw. Spreizklimmern (Lonicera implexa, Rubia peregrina, Asparagus albus) als zusätzlichem Strukturelement (J43).
Immergrüne Eichen-Arten sind hier nicht wettbewerbsfähig. Lokal kann
der mittlere Jahresniederschlag bis auf ca. 200 mm sinken bzw. der
Standort edaphisch (Gipsmergel) oder durch Windeinwirkung zusätzlich
physiologisch trocken sein. Hier ist auch dem Pistaziengebüsch die
Existenzgrundlage entzogen, und es treten semiarid-aride Trockengebüsche
mit dominierendem Ziziphus lotus an seine Stelle (J52). In den trockensten und windigsten Küstenlagen (Cabo de Gata bis Carboneras) ist Periploca angustifolia (J53), im Gebiet um Almeria Maytenus senegalensis subsp. europaea die Leitart der Trockengebüsche (J51). Die mesomediterrane Stufe des gebirgigen Hinterlandes wird bei mittleren jährlichen Niederschlagsmengen von 400-500 mm von Juniperus oxycedrus-reichen Kermeseichengebüschen (Quercus coccifera) – oft mit Pinus halepensis-Überschirmung – eingenommen. Die Ausbildungsform im Bergland nördlich von Almeria (J31) ist durch geographische Differentialarten (schwach) von der im Bergland oberhalb von Murcia (J30) abgrenzbar. Erst mit zunehmenden Niederschlägen in supramediterranen Lagen (ca. 900-1900 m) wird Quercus ilex subsp. rotundifolia wettbewerbsfähig und überlagert östlich der Sierra Nevada die Quercus coccifera-Stufe sowohl auf Kalksubstraten (J9, mit Juniperus thurifera) als auch über Silikat- und Serpentingesteinen (J7, mit Adenocarpus decorticans).
Das
kontinental getönte I n n e r e d e r I b e r i s c h e n Ha l b i n s e
l (Castilla - La Mancha, Extremadura) wird großflächig von
mesomediterranen Quercus ilex subsp. rotundifolia-Wäldern eingenommen.
Kernwüchsige Altbäume sind vor allem in der Extremadura als Mastbäume in
offenen Hutewäldern (dehesas) vielerorts erhalten geblieben. Die über
Silikatgesteinen weit verbreitete Ausbildungsform dieser Wälder (Pyro bourgaeanae-Quercetum rotundifoliae, J1)
ist durch entsprechende edaphische Zeigerarten differenziert von einem
basiphilen Flügel, der ein westliches Teilareal in der Extremadura (Paeonio coriaceae-Quercetum rotundifoliae, J2) und ein östliches Teilareal in Neukastilien (Bupleuro rigidi-Quercetum rotundifoliae,
J4) besiedelt. Nördlich des Kastilischen Scheidegebirges ist auf der
altkastilischen Hochfläche von Salamanca und Valladolid die
Differenzierung der Quercus ilex subsp. rotundifolia-Wälder
vergleichbar: Die Bestände der dortigen Silikat- und Serpentingebiete
sind durch das Vorhandensein bzw. Fehlen ozeanisch getönter
Differentialarten in einen westlichen Flügel (Genisto hystricis-Quercetum rotundifoliae, J5) und einen östlichen Flügel geschieden (Junipero oxycedri-Quercetum rotundifoliae, J6), denen die basiphilen Ausbildungen der mesozoischen und tertiären Kalksteinlandschaften gegenüberstehen (Junipero thuriferae-Quercetum rotundifoliae, J9). Nördlich von Burgos klingen die inneriberischen Quercus ilex subsp. rotundifolia-Wälder
an der Südabdachung der Kantabrischen Kordillere aus, supramediterran
beschränkt auf edaphische Sonderstandorte über kompaktem Kalkgestein (Spiraeo obovatae-Quercetum rotundifoliae, J8).
Die genannten mittel- und ostiberischen Quercus ilex subsp.
rotundifolia-Wälder kleiden auch – in gleicher Weise nach Substrat und
Höhenlage differenziert – die mesomediterrane Gebirgsumrahmung des
nordostiberischen Ebro-Beckens im Hinterland von K a t a l o n i e n
aus. Das küstenferne Innere des Beckens um Zaragoza wird hingegen auf
Grund der geringen mittleren Jahresniederschläge von mesomediterranen
Kermeseichen-Buschwäldern eingenommen (Rhamno lycioidis-Quercetum cocciferae, J30). Die Kermeseiche ist auch in den thermomediterranen Quercus ilex subsp. rotundifolia-Wäldern
an der ostspanischen Küste zwischen Valencia und Tarragona als
diagnostisch und strukturell wichtige Begleit-Holzart stärker beteiligt (J39); sie kann in semiariden Situationen im Gebiet zwischen Valencia und Alicante auch mit Pistacia lentiscus Küstengebüsche bilden (J44), in denen Quercus ilex subsp. rotundifolia
nicht konkurrieren kann und die südwärts zu dem bereits genannten
Pistazienbusch des südostspanischen Trockengebietes überleiten.
Nordwärts hingegen klingen die katalonischen Quercus ilex subsp. rotundifolia-Wälder am Südfuß der Pyrenäen in Form reliktischer Bestände mit reichlich Buxus sempervirens aus (Helleboro foetidi-Quercetum rotundifoliae, J12). Im nördlichen Abschnitt der katalonischen Küste bei Barcelona, d. H. außerhalb des Areals von Quercus ilex subsp. rotundifolia, treten unter thermisch ausgeglicheneren „nordmediterranen“ Klimabedingungen schließlich Quercus ilex subsp. ilex (J14) und auch wieder Quercus suber (J27) als Waldbildner in Erscheinung.
Die
der katalonischen Küste vorgelagerten B a l e a r e n werden ganz von
immergrüner Hartlaubvegetation eingenommen, die floristisch durch
Inselendemiten von den festländischen Ausbildungsformen Spaniens
synchorologisch abgrenzbar ist. Auf Mallorca zeigt sich eine weitere
Differenzierung im wesentlichen nach thermischen Parametern. Die
südliche Küstenregion ist als Wuchsgebiet trockenresistenter
Johannisbrotbaumwälder ausgewiesen (J46), das weniger trockene Innere und die Nordküste als Wuchsgebiet des thermomediterranen Quercus ilex subsp. rotundifolia-Waldes (J40).
Die
Gipfellagen des Inselinneren tragen dagegen bei ausreichendem mittlerem
Jahresniederschlag einen mesomediterranen Steineichenwald (Cyclamini balearici-Quercetum ilicis, J16). Insuläre Isolationseffekte bedingen floristische Unterschiede in der Thermomediterran-Vegetation der Nachbarinseln Menorca (J47) und Ibiza (J45).
Das
mediterrane S ü d f r a n k r e i c h ist wissenschaftsgeschichtlich
mit der pflanzensoziologischen Erforschung der immergrünen
Hartlaubvegetation Südeuropas eng verbunden. Hier entwickelte
Braun-Blanquet sein Konzept vom geschlossenen Steineichenwald (mit Quercus ilex subsp. ilex) als Reifestadium der natürlichen Vegetation des mediterranen Winterregengebietes („Quercetum ilicis gallo-provinciale“
Br.-Bl. 1936) und erhellte die syndynamischen Zusammenhänge zur
aktuellen Sekundärvegetation der Macchien und Garrigues. Tatsächlich
geschah dieses eigentlich am floristisch „verarmten“ Nordrand der
Mediterranvegetation, wo die thermomediterrane Zone bzw. Stufe aus
klimatischen Gründen nahezu ganz ausfällt (bis auf inselartige Vorkommen
bei Monaco,
J48).
Potentiell besiedeln mesomediterrane Quercus ilex-Wälder die
küstennahen Gebiete Nordkataloniens, des Languedoc, der Provence und der
französischen Riviera (Viburno tini-Quercetum ilicis).
Kennzeichnend (wenn auch keineswegs syntaxonomisch und synchorologisch
auf sie beschränkt) ist ihr Reichtum an Lianen und Spreizklimmern (Smilax
aspera, Rubia peregrina, Tamus communis, Lonicera implexa, Rosa
sempervirens, Asparagus acutifolius, Clematis flammula, Hedera helix). Auf flachgründigen Böden und als Störungszeiger können Pinus pinaster und P. halepensis
in der Baumschicht eine Rolle als Begleit-Holzarten spielen. Am Ostfuß
der Pyrenäen (Roussillon) sowie an der französischen Rivieraküste
zwischen Toulon und Cannes (Massif de Maures) ist auf oligotrophen,
lessivierten Silikatböden dagegen die Korkeiche der Hauptwaldbildner (Carici depressae-Quercetum suberis, J27).
Im küstenferneren, mittelgebirgigen Hinterland Nordkataloniens
(Montserrat-Massiv) und Südfrankreichs (im Rhonetal nordwärts bis
Valence) stockt ein supramediterraner Steineichenwald (Asplenio onopteridis-Quercetum ilicis, J15), das klassische „Quercetum ilicis mediterraneomontanum“
Br.-Bl. 1936, in welchem immergrüne Begleit-Holzarten mit zunehmender
Küstenferne zurücktreten und winterkahle Gehölze, vornehmlich Quercus
pubescens, ihre Stelle einnehmen.
Formation J (Subformationen J.1, J.2) auf der Iberischen halbinsel
| |
 |  |
Wichtige, an die Serra da Estrela angrenzende Kartierungseinheiten der Formation J

J5 - Old Castilian supra-Mediterranean Quercus ilex subsp. rotundifolia-forests with Genista hystrix on siliceous and serpentine rocks

J25 - Southwest Iberian meso-Mediterranean cork oak forests (Quercus suber) with Sanguisorba hybrida, Paeonia broteroi
Fortsetzung -> Teil IV
Fortsetzung -> Teil IV