Eine Botanisch-Zoologische Rundreise auf der Iberischen Halbinsel.
Auf der Suche nach der Verlorenen Zeit
Von Horst Engels
Teil II - Reisestationen

Reisestationen der Botanisch-Zoologischen Rundreise um die Iberische Halbinsel
2.5 Die Nördlichen Gebirge Portugals (Northern Serras of Portugal)
2.5 Die Nördlichen Gebirge Portugals
2.5.2 Serra da Estrela
|
Tabellenkalkulation:
|
Datenbanken:
|
Kartenmaterial:
|
2.5.2 Habitate und Gemeinschaften in der Serra da Estrela
C5. Die Felsumgebung der Serra da Estrela

Rupikole Vegetation im Basalen Höhengürtel (500-800 m NN) der Serra da Estrela (Moinhos de Aveia, Pinhel)

Flechten, Moose und Gräser auf Granitfelsen und in deren Spalten im Montanen Höhengürtel (800-1600 m NN) der Serra da Estrela (oberhalb Lagoa Comprida)


Gelber Enzian (Gentiana lutea) - im hohen (?oro-mediterranen - ?oro-temperaten) Gürtel (1600-1993 m NN) der Serra da Estrela

Saxifraga spathularis (Spatelblättriger Steinbrech) -
Felsspaltenvegetation im hohen Gürtel (1600-1993 m NN) der Serra da Estrela
Felshabitate (Natura 2000)
Felshabitate finden sich an zwei ganz unterschiedlichen Orten, zum einen an den Meeresküsten, wo Felsklippen und Riffe auftreten, zum anderen in den Gebirgen. In den Hochgebirgen überwiegen sogar die rohen Felsoberflächen, aus denen durch Metereorisierung Geröllhalden und durch Erosion und Humusanreicherung die Böden der Ebenen hervorgehen. Je nach Art des Gesteins, ob basisch oder sauer, finden sich in den Felsumgebungen die unterschiedlichsten Vegetationstypen. Auch die Schutthalden besitzen ihre eigene Vegetation. Natürlichen Höhlen und Grotten werden ebenfalls als Felsumgebung eingestuft.
In Natura 2000 sind die folgenden Felshabitate für Portugal eingetragen:
8 Felshabitate und Höhlen
81 Geröllhalden an Felshängen
8130 Westmediterrane und thermophile Geröllhalden
82 Felshänge mit chasmophytischer Vegetation
8210 Felshänge mit chasmophytischer Vegetation auf Kalkgestein
8220 Felshänge mit chasmophytischer Vegetation auf Silikatgestein
8230 Silikatfelsen mit Pioniervegetation des Sedo-Scleranthion oder des Sedo albi-Veronicion dillenii
8240 * Kalklapias
83 Andere Fels-Habitate
8310 Vom Tourismus nicht explorierte Höhlen
8330 Submerse oder semi-submerse Meeresgrotten
Für die Serra da Estrela (mit überwiegend silikatreichem Gestein) sind von Bedeutung:
Habitate in Geröllhalden (engl. screes) - westmediterrane und thermophile Geröllhalden (Habitat 8130pt2); Hänge mit Silikatgestein (Habitat 8220); Silikatfelsen mit Pioniervegetation des Sedo-Scleranthion ou da Sedo albi-Veronicion dillenii (Habitat 8230pt2); Habitate der von Touristen nicht besuchten Grotten (Habitat 8310).
Geobotanische Charakterisierung der Felsumgebungen der Serra da Estrela (Nach Jan JANSEN, 2002)
Jan JANSEN (2002) charakterisiert die ökologischen und geobotanischen Eigenschaften der estrelensischen Felsumgebungen im “Geobotanical guide of the Serra da Estrela” (pp. 175-192):
Organismen, die auf Felsen leben, können Wasser nur aus direktem Niederschlag, Abfluss oder Versickern erhalten. Organismen, die ausschließlich an atmosphärisches Wasser gebunden sind, sollten deshalb in der Lage sein, auftretende Intervalle ohne jede Wasserversorgung zu überleben. Alle von ihnen sind wechselfeucht (poikilohydrisch), was bedeutet, dass sie sich in einem Ruhezustand am Leben erhalten, wenn sie völlig von der Wasserversorgung ausgeschlossen sind. Die meisten von ihnen sollten auch an ein extremes Temperaturregime angepasst sein, da die Felsoberflächen oftmals über kürzere oder längere Intervalle großen Temperaturschwankungen ausgesetzt sind, wodurch auch der Wassermangel verstärkt wird. Im oberen Gürtel können Temperaturextreme auf Felsoberflächen von weniger als -10 °C bis von über +50 °C auftreten. Gefäßpflanzen können nicht auf nackten Felsoberflächen wachsen, aber einige Algen, Moose (Bryophyten) und besonders viele Flechten können trotz der extremen Umweltbedingungen auf Felsen wachsen. Eine Flechte ist die symbiotische Verbindung eines Pilzes und einer Alge. Eine Symbiose ist die Interaktion zwischen zwei und mehr Arten, die mindestens einer von ihnen profitiert. Nur bestimmte Pilze und Algen können zusammenkommen, um einen Flechtenkörper zu bilden. Die Alge benutzt Sonnenlicht, um Zucker zu erzeugen, die sowohl Pilz als auch Alge ernähren wird. Der Pilz schafft seinerseits einen Körper, der beide Organismen beherbergen kann. Es gibt mehrere Arten von Flechten. Krustenähnliche Flechten haften fest an den Felsen an ihrer ganzen Unterseite und manche können sogar in den Felsen eingebettet werden. Blattähnliche Flechten haben eine ausgeprägte obere und untere Oberfläche und werden nur durch kleine wurzelartige Strukturen befestigt. Schließlich bilden strauchähnliche Flechten verästelte Stränge oder Hohlstiele und sind gewöhnlich an der Basis befestigt. Die meisten der exponierten Felsen in der Gegend sind weitgehend von Flechten bedeckt. Der obere Gürtel des Estrela beherbergt etwa 250 Flechtenarten, etwa etwas mehr als die Hälfte von ihnen können auf nackten Felsen wachsen. Diese Arten können in verschiedenen Assemblagen auf Felsoberflächen in typischen Felsumgebungen wie Klippen, Tors, Schutthalden und felsigen Süßwasser-Flussbetten auftreten. Die Flechten- und Bryophyten-Gemeinschaften können hier nicht behandelt werden. Darüber hinaus sind die meisten von ihnen noch nicht untersucht worden. Fast alle Bryophyten und Gefäßpflanzen brauchen mindestens eine feine Erdschicht, die aus gefallenen Partikeln oder eingeblasenem Staub entsteht.
Felsbewohnende (rupikole oder saxikole) Arten wachsen in felsigen Umgebungen. Saxikol (engl. saxicolous) kommt aus dem lateinischen 'saxum' (rock) und 'colere' (bewohnen); rupikol (engl. rupicolous) ist eine Übersetzung von 'rupestris' (was auf Felsen lebt). Die Spezies, die auf Felsoberflächen wachsen, können als Chomophyten (griech. chõma »aufgeschüttete Erde«) (Detrituspflanzen), die in Felsspalten wachsenden als Chasmophyten (lat. chasma »Riss, Kluft«) (Felskluftpflanzen) bezeichnet werden. In der Praxis kann man an beiden Standorten eine beträchtliche Anzahl von rupikolen Arten finden.
Eine vorläufige Klassifikation der rupikolen Pflanzen aufgrund ihrer Biotop-Präferenzen ("first choice biotopes") kann nützlich sein, um ein besseres Verständnis für ihre optimalen Wachstumsbedingungen zu erhalten. Hier werden zwei Hauptgruppen von rupikolen Spezies unterschieden, eine mit Vorliebe für Gesteine (Fissuren, Gesteinsoberflächen und felsige Hänge), die andere mit Vorliebe für Schutt- oder Geröllhalden (Jan JANSEN, 2002, pp. 175-193).
Felsspalten, Felsoberflächen und Talushänge
In diesem Abschnitt werden auffällige Felsspezialisten unter den Gefäßpflanzen und ihre Umgebung eher global beschrieben, da sich die Umweltbedingungen über sehr kurze Distanzen ändern können, wodurch eine Unterscheidung bestimmter Gemeinschaften eher willkürlich wird. Bevor wir diese Felsbewohner beschreiben, muss noch erwähnt werden, dass eine Diagnose ihrer Gemeinschaften oft dadurch erschwert wird, dass sie oft in der Nähe von Arten wachsen, die für andere Biotope charakteristisch sind. Elemente der natürlichen rupikolen Vegetation können sich in künstlichen anthropogen erzeugten Biotopen wie Mauern und Straßenrändern zeigen. Da diese Orte oft durch Nährstoffe angereichert sind, können sich (sub-)nitrophilie-Arten anschließen und die Vorherrschaft gewinnen. Darüber hinaus treten Talushänge und Waldränder oft eng miteinander auf, und ihre Pflanzengemeinschaften können sich vermischen, so dass eine klare Trennung unmöglich wird. In gleicher Weise sind viele einjährige Grasland-Spezies in der Lage, erodierte Felsoberflächen und Felsspalten zu kolonisieren, sobald sie eine minimale Erdschicht enthalten.
Charakteristische rupikole Pflanzen der Serra da Estrela sind:
 |  |
 |  |
 |  |
 |  |
  | 
Narcissus rupicola
|
  |  |
 |   |
 |  |
 |  |
 |  |
  |  |
  |  |
 |  |
 |  |
 |  |
 |  |
 |  |
1.1 Basale Höhenzone (Basaler Gürtel) 500-800m NN
Entlang einiger Talhänge im Basalen Gürtel kann eine seltene Farngemeinschaft mit Billots Streifenfarn (Asplenium billotii) und einer Pelzfarnspezies (Cheilanthes tinaei der Gattung Cheilanthes (die weltweit ca mit 180 Arten und in Portugal mit 5 Arten vertreten ist) in Spalten schistöser Felsaufschlüsse auftreten. Risse und Fissuren der gleichen Art von Muttergestein, aber meist in höheren Höhen, können kleine Populationen von Murbeckiella sousae, einem Mitglied der Familie der Kreuzblütler (Brassicaceae), beherbergen. Exponierte Klippen in der Nähe von windgepeitschten Kämmen und Gipfeln scheinen optimale Bedingungen für diese Art, die streng endemisch für Portugal ist, zu liefern. Die Art ist im Rahmen der EU-Habitatrichtlinie (Anhang IV) geschützt. Ihr Verbreitungsspektrum umfasst die unteren und mittleren Gürtel einiger Gebirge in Zentral- und Nord-Portugal, so die der Serra da Lousã, Serra do Açor, Serra da Estrela, Serra da Freita, Serra do Alvão und Serra do Marão.
Murbeckiella boryi, ein enger Verwandter, der innerhalb Portugals auf die Estrela beschränkt ist (ein weiterer Nachweis ist bisher nur für die Serra do Alvão in der Flora-On publiziert), wächst im oberen Gürtel in granitischen Felsfissuren. Neben den verschiedenen Lebensraumpräferenzen kann Murbeckiella sousae (unter anderen morphologischen Merkmalen) durch ihre kleineren Blütenblätter (3,5-4 mm) und ihre graugrünen Blätter von dieser Art unterschieden werden. Blütenblätter von Murbeckiella boryi besitzen die Maße 3-7 (8) mm und ihre Blätter sind dunkelgrün.
Sonnige Felsnasen, Wände und Felsablagerungen, aus Granit oder metasedimentärem Fels entstanden, können auffällige Arten wie Digitalis thapsi, Dianthus lusitanus, Rumex induratus und Phagnalon saxatile beherbergen. Alle vier Arten treten im unteren und mittleren Gürtel auf, aber Dianthus lusitanus ist die einzige Spezies, die in den hohen Gürtel vordringt. Diese rosa-blühende iberische endemische Nelkenspezies ist offensichtlich in der Lage, große Temperaturextreme zu bewältigen. In dieser Umgebung kann Dianthus lusitanus Mosaike mit Polsterkissen-Gebüsch bilden.
Weniger xerische Felsumgebungen können von dem Leimkraut Silene acutifolia kolonisiert werden. Dieser rosafarbene, blühende Felsbewohner ist im mittleren (montanen) Gürtel relativ verbreitet, aber dringt auch bis in die höchste Zone vor, wo die Art bisweilen mit Silene x montistellensis vergesellschaftet ist oder durch diese ersetzt wird. Silene acutifolia kolonisiert sowohl Felsspalten als auch flache Skelettböden aus Granit oder Metasedimenten. Sie scheint höhere Nährstoffanforderungen zu haben als die beiden kleineren Murbeckiella Arten. Im mittleren Gürtel kann sie in der Nähe von Murbeckiella sousae Populationen auftreten, die von kleineren Rissen profitieren können, vor allem bei überhängenden Aufschlüssen. Auf steilen Klippen im Oberen Gürtel kann sie in breiteren Fissuren neben kleineren Rissen mit Murbeckiella boryi wachsen, Silene acutifolia kann mit Silene foetida verwechselt werden, mit dem sie auch hybridisiert. Allerdings wächst S. acutifolia in Felsspalten und S. foetida auf den Schutthalden. Sie können morphologisch wie folgt unterschieden werden: S. acutifolia hat eine basale Blattrosette, einen aufgerichteten Stengel und ganze oder fast ganze rosa purpurrote Blütenblätter: S. foetida besitzt keine Basalblattrosette, einen Stengel, der lose an der Bodenoberfläche liegt und tief eingekerbte blassrosa oder weiße Blütenblätter. Stengell und Blattrosette der Hybrid S. x montistellensis sind ähnlich wie S. foetida, ihre Blütenblätter sind weniger tief eingekerbt, aber die Farbe der Blütenblätter ist ähnlich wie S. acutifolia. S. foetida und die Hybridform sind auf die Estrela beschränkt. S. acutifolia hat eine westiberisches Verbreitung, welche die Gebirgsregionen des Alto Alentejo, sämtlichen Beira-Provinzen, Douro Littoral, Minho und Tras-os-Montes in Portugal einschließt. Sie kommt auch in Orense und Lugo in Spanien vor.
1.2 Basaler und Mittlerer (Montaner) Gürtel - 800-1600m NN

Silene acutifolia, Covão da Ametade (Serra da Estrela)
Im unteren und mittleren Gürtel, vor allem in den östlichen Parkgebieten mit einem weniger atlantischen Klima, jedoch vielleicht unter lokal feuchteren Wachstumsbedingungen, kann Silene acutifolia durch Saxifraga fragosoi ersetzt werden. Das Verbreitungsgebiet von Saxifraga fragosoi erstreckt sich weniger westlich und südlich, aber weiter in nördlicher und östlicher Richtung, darunter in die portugiesischen Provinzen Beira Alta, Beira Baixa und Tras-os-Montes und einen großen Teil der nordspanischen Provinzen, die fast bis an die Pyrenäen reichen. Außerhalb der Iberischen Halbinsel wächst sie in den Gebirgen Südfrankreichs. Die weissblühenden Teppiche von Saxifraga fragosoi sind vom späten Frühjahr bis zum Frühsommer, vor allem auf verhältnismäßig steilen, meist nordwestlich bis nordöstlich exponierten Felshängen, zu sehen. Nach dieser Zeit fallen die sonnenverwöhnten Teppiche besonders durch ihre rötliche Farbe auf.

Saxifraga fragosoi -Teppich (rotbraun) im Covão da Ametade, Serra da Estrela

Saxifraga fragosoi - Covão da Ametade, Serra da Estrela

Saxifraga fragosoi - Covão da Ametade, Serra da Estrela
Saxifraga fragosoi Vegetation kann im unteren Gürtel auftreten, erreicht aber seine optimale Entwicklung im mittleren Gürtel. Hier nimmt es flache Skelettböden ein und kann die Felsoberflächen (Granit, Metasediment), oft neben Pflanzengesellschaften der tiefgründigeren Böden wie Waldrand-, Busch- und Graslandgesellschaften abdecken. Saxifraga fragosoi ist häufig mit Phalacrocarpum oppositifolium, Sedum hirsutum und Sedum brevifolium vergesellschaftet. Gelegentlich erscheinen auch andere Mauerpfeffer (Fetthenne) wie Sedum anglicum subsp. pyrenaicum, Sedum forsterianum und Sedum pruinatum.
Mauerpfeffer (Sedum) ist eine Gattung aus der Dickblatt-Familie (Crassulaceae). Mitglieder dieser Familie besitzen CAM-Metabolismus. Dies ist eine photosynthetische Strategie, die für Sukkulenten charakteristisch ist, bei der CO2 nachts chemisch "fixiert" wird und während des Tages freigelassen wird, wenn die Spaltöffnungen (Stomata) geschlossen bleiben. Dadurch wird die innere Feuchtigkeit bewahrt. Dieser Mechanismus ist eine hervorragende Anpassung an xerische Biotope wie Felsen. Die meisten Mauerpfeffer-Arten scheinen sich optimal auf flachen oder sanft abfallenden Substraten zu entwickeln und treten häufig zusammen mit andere Formationen wie Grasland und Gebüschen auf, wenn auch in kleinen Zahl von Spezies.
Im unteren und mittleren Gürtel kann Sedum pruinatum auf sanft geneigten Talushängen unter steilen Klippen, die zum Teil mit Murbeckiella sousae und Silene acutifolia bewachsen sind, Matten bilden. Sedum pruinatum ist ein weiteres west-iberisches endemisches Taxon der Felsbiotope. Es fehlt in den Provinzen Alto Alentejo, Beira Litoral und Luso, aber sonst ist seine Verbreitung ähnlich wie die von Silene acutifolia. Das gelbblütige Sedum pruinatum ist in der Lage, lose verwobene Zöpfe auf sanft abfallenden Felsen und schlecht entwickelten Böden zu bilden, oft an exponierten und nordorientierten Standorten. Sowohl auf Metasedimenten als auch auf Graniten ist es oft mit Arten vergesellschaftet, die für offene Wiesen oder niedrige Gebüsche charakteristisch sind. Die Bestände der Estrela werden häufiger von Orophyten als Sedum pruinatum Gesellschaften aus Nordportugal und Galizien begleitet.
In der Estrela kommen mindestens drei einjährige Mauerpfeffer-Arten vor. Aufgrund ihrer Lebensform werden sie oft als Charakterarten von therophytischen Formationen betrachtet: Sedum arenarium und Sedum pedicellatum subsp. lusitanicum von therophytischen Rasen und Sedum maireanum von saisonal überschwemmten Biotopen. Allerdings besitzen alle drei Vorlieben für Kies und felsige Umgebungen. Sie können leicht übersehen werden, außer den häufig rot gefärbten Flecken von Sedum arenarium, die sanft abfallende felsige Substrate in niedrigeren und mittleren Erhebungen im Gebiet dominiert.
Eine der häufigsten Sedum-Arten ist Sedum brevifolium, obwohl sie selten den Bestand dominiert. Sie ist fähig, felsige Lebensräume und Kies zu kolonisieren und kann in verschiedenen Formationen wie Wiesen und Sträuchern bestehen bleiben. Es ist in niedrigeren Höhen zu finden, aber am besten in den höchsten Teilen des Berges entwickelt. In relativ feuchten Felsvorsprüngen wird oft zusammen mit Sedum anglicum subsp. pyrenaicum angetroffen. An diesen Orten kann letztere dominant werden, vor allem unter hohen Klippen in flachen Kiesböden, meist in der Nachbarschaft von besser entwickelten Böden mit Festuca henriquesii Rasen.
 | 
Sedum brevifolium
|
 |  |
 |  |
 |  |
1.3 Oberer Gürtel (Höhenstufe von 1600-1993 m NN)
Das Bioklima des Obersten Höhengürtels der Serra da Estrela wird von einigen als oro-temperat (alpin), von anderen als oro-mediterran (subalpin) eingestuft, weshalb auch unterschiedliche Ansichten über die Zuordnung des Sektors ‘Estrelense’ der Serra da Estrela zu einer mediterranen oder einer gemäßigt atlantischen Florenprovinz bestehen.
Jan JANSEN (2002) geht von einem oro-temperaten Bioklima im höchsten Gürtel aus. Von anderen Autoren, z..B. BOHN et al. (2002), wird diese von Polunin & Smythies und auch von Jan JANSEN (2002) als alpin (oro-temperat) bezeichnete Zone der Serra da Estrela als oro-mediterran (subalpin), und damit nicht als eine natürlich baumfreie Zone zwischen der oberen Baumgrenze und der permanenten Schneezone (nivale Stufe) angesehen. Es herrscht also weiterhin Uneinigkeit darüber, ob eine natürliche Baumgrenze, und damit eine baumfreie, alpine Zone in der Serra da Estrela existiert oder ob die Hochebene der Serra da Estrela früher bewaldet war und lediglich eine anthropogene Abholzung zu der jetzt baumfreien Zone der Serra da Estrela geführt hat. MALKMUS (1985) sieht die Möglichkeit der Existenz einer früheren Bewaldung der Hochebene aufgrund der rezenten Herpetofauna. Auch Pollenanalysen sprechen für eine frühere Bewaldung.
Hier zunächst einige Fotos von Pflanzen der Felsumgebungen dieser höchstgelegenen Zone der Serra da Estrela, bevor wir auf ihre Beschreibung durch Jan JANSEN (2002) zurückkommen:

Senecio pyrenaicus subsp. caespitosus





Gentiana lutea subsp. lutea (?aurantiaca) existiert in Portugal nur im obersten Gürtel der Serra da Estrela, wo die Art in den Rasen der Kare und Felsumgebungen auftritt. Es ist die unzugängliche Felsumgebung der Cântaros und die der aus der Eiszeit verbliebenen Kare dieses Hochgebirges,, die das Überleben dieser Art in der Serra da Estrela ermöglichen. Denn die Wurzeln von Gentiana lutea werden aufgrund ihrer phytotherapeutische medizinischen Eigenschaften gesammelt, weshalb das Überleben dieser Art in der Serra da Estrela stark gefährdet ist.


Jasione spec.








In Schattige Felsspalten im oberen Gürtel fallen die fleischigen Rosetten des Spathelblättrigen Steinbrechs (Saxifraga spathularis) auf. Diese Art ist am häufigsten im Oberen Gürtel anzutreffen, kann aber auch auf niedrigerer Höhe in Birkenwäldern und in der Nähe von Quellen vorkommen. Saxifraga spathularis bevorzugt ziemlich feuchte Fissuren; sie wird oft in Gesellschaft von von Murbeckiella boryi angetroffen, obwohl letztere Art häufiger in trockeneren Rissen und Fissuren auftritt. Nur an wenigen schattigen Plätzen findet sich der seltene Frauenmantel Alchemilla transiens, eine Staude aus der Familie der Rosengewächse (Rosaceae) mit dichten Clustern von winzigen grünlich-gelben Blüten. In breiteren, mit Boden gefüllten Felsspalten an schattigen Orten werden die vorgenannten Arten durch Spezies der Festuca henriquesii-Rasen oder unter exponierteren Bedingungen durch Arten von xerophytischen mehrjährigen Graslandgesellschaften begleitet oder ersetzt.
Besonders in überhängenden Granitfelsen können sich Farne wie Schwarzstieliger Streifenfarn (Asplenium adiantum-nigrum), Braunstieliger Streifenfarn (Asplenium trichomanes subsp. quadrivalens), Rippenfarn (Blechnum spicant) und Nordischer Streifenfarn (Asplenium septentrionale) verbergen. Letzterer tritt in großen Teilen der nördlichen Hemisphäre auf, ist aber in Portugal und außerhalb von Estrela nur von wenigen Orten um Braganca und Vinhais bekannt. Andere Farne in großen Höhen des Oberen Gürtels sind der Krause Rollfarn (Cryptogramma crispa) und der Wurmfarn Dryopteris oreades. Sie können Risse und Fissuren der Felswände kolonisieren, aber die Chance ist größer, sie zwischen großen Felsbrocken in den Geröllhalden zu finden.
2. Geröllhalden (engl. screes) des Oberen Gürtels (1600-1993 m NN)
(Ordnungen Androsacetalia alpinae und Polystichetalia lonchitidis (Klasse Thlaspietea rotundifolii))
Otti WILMANNS (1998) schreibt ausführlich zu den Standortfaktoren von Gesteinsschuttfluren der Alpen und übrigen westlichen Hemisphäre:
Edaphische Standortsfaktoren
Eine bewegte – und damit in ihrer Vegetation typische – Schutthalde ist ein Ökosystem in offenkundigem Fließgleichgewicht. Durch Schwerkraft, Wasser- und/oder Eistransport kollern und rutschen Gesteinsbrocken verschiedenster Größe von anstehenden Felswänden herab und bilden die Halde. (Unter Schutt im geologischen Sinne werden kantige Steine beliebiger Größe verstanden, während Schotter (3-30 cm Ø) vom Wasser transportiert und dabei gerundet worden sind.) Ist ein vom Schuttmaterial abhängiger Neigungswinkel (etwa 30-35°) erreicht, so kann das instabile Gleichgewicht durch frischen Schutt, durch Wildtritt, durch Schneedruck oder auf ähnliche Weise leicht gestört werden; die Schutthalde bleibt in Bewegung.
In diesem Falle bleiben die Pioniergesellschaften über Jahrtausende erhalten; sie sind zugleich Dauergesellschaften. Eine Sukzession über Folge- zu Schlußgesellschaften kann nur dann stattfinden, wenn sich die physikalischen Standortsbedingungen ändern, wie es z. B. im Schuttfeld vor abschmelzenden Gletschern geschieht (s. u.).
Die Besonderheit der Schutthalde als Extremstandort wird erst recht klar, wenn man ein Bodenprofil ansieht: Auf eine obere sog. Stein-Luft-Schicht wechselnde Mächtigkeit, i. a. etwa 10-30 cm, folgt eine Feinerde-Stein-Schicht: diese liegt einer unteren Stein-Luft-Schicht auf; nur letztere ist unbewegt. Entscheidend ist die Ausbildung der Feinerde-führenden Schicht. Diese besteht aus eingewaschenem Staub und aus Pflanzenresten. BRAUN-BLANQUET & JENNY (1926) bestimmten im Schweizerischen Nationalpark 1,4-1,85 kg Flugstaubniederschlag pro m2 und Jahr. Er sammelt sich an Stellen, wo die Steine dicht lagern; hier kann langsam und spärlich Humus gebildet werden. An diesen Rohböden, die, wenn jenseits der Waldgrenze liegend, Råmark, innerhalb des potentiellen Waldgebietes Syrosem genannt werden, spielt sich die weitere Bodenbildung ab.
Die Verwitterungsweise des Muttergesteins wirkt sich bestimmend auf die Vegetation aus; aus dem BS 8 geht hervor, daß nicht nur die üblichen Haupttypen Kalk- und Silikatgestein geschieden sind, sondern innerhalb des letzteren die in den Zentralalpen anstehenden Kalkschiefer eigene Gesellschaften tragen (ZOLLITSCH 1966). Die unterschiedlichen ökologisch wirksamen Bodenfaktoren der beiden Haupttypen sind auf S. 206 zusammengestellt. Die Gründe für die Eigenständigkeit der Kalkschiefer-Vegetation sind trotz der Bodenanalysen von ZOLLITSCH noch nicht klar; die pH-Werte, um 8 liegend, ähneln denen der Kalkschutthalden, ihr Feinerdereichtum scheint dem von Silikatschutt- und Mergelhalden zu entsprechen; die Humusgehalte überlappen sich. Offenbar gibt es noch unerkannte differenzierende, qualitative Merkmale.
Den Assoziationen der Epilobietalia fleischeri ist gemeinsam, daß sie auf grobem, feinerdearmem, oberflächlich rasch und stark austrocknendem Substrat leben (Abb. 26). Aufgefüllt wird das Wasserreservoir durch ein- oder gar mehrmalige Überschwemmungen während der Vegetationsperiode, denn sie besiedeln ja Flußbetten, welche die sommerlichen Schneeschmelzwässer der Alpen abführen. Deren Gewalt reißt auch die Kies- und Sandbänke immer wieder fort und lagert sie um, so daß die Sukzession vielfach abgebrochen wird. Hier sind denn auch die chemischen Substratunterschiede ausgeglichen.
Klimatische Standortsfaktoren
An dieser Stelle sollen ganz knapp wesentliche Züge des Pflanzenklimas in der alpinnivalen Stufe der Alpen, die auch auf andere Hochgebirgsgesellschaften übertragbar sind, erläutert werden, denn die aufschlußreichen, mit modernster Meßtechnik durchgeführten ökologischen Untersuchungen der „Innsbrucker Schule" im Rahmen des Internationalen Biologischen Programms am Hohen Nebelkogel in 3184 m bezogen sich überwiegend auf Pflanzen der Androsacetalia alpinae (MOSER et al. 1977, Zusammenfassungen bei LARCHER 1980, 1994a). Zu diesen gehören auch die höchststeigenden Pflanzen der Schweizer Alpen, welche die Schneegrenze noch um mehr als 600 m unter sich lassen: Androsace alpina, Ranunculus glacialis, Saxifraga bryoides, S. exarata (auch im Caricetum curvulae) und S. oppositifolia (Thlaspietea).
Zwei Tatsachen sind grundlegend für das Verständnis der klimatischen Eigenart der Hochgebirgslagen:
1. Die Luft ist „dünner" und „reiner", d. h. sie besitzt weniger Masse pro Volumeneinheit und sie enthält weniger Wasserdampf und Staub. Das bewirkt (gemäß den Gesetzmäßigkeiten des Strahlungsumsatzes an der Erdoberfläche), daß die Einstrahlung bei Tag und die Ausstrahlung bei Nacht stärker sind als in den Tieflagen; ferner laufen Erwärmung und Abkühlung der Luft rascher ab.
2. Die Windgeschwindigkeiten sind höher. Daraus ergeben sich für die Lebewesen Existenz-bestimmende Standortseigenschaften, die vielfach noch durch die Steilheit der Hänge verstärkt werden:
- Es kommt tagsüber zu starker Erwärmung des Bodens und der Pflanzen.
- Nachts ist die Abkühlung beider besonders ausgeprägt; selbst im Sommer können Fröste auftreten, die bis in 10 cm Tiefe reichen.
- Es gibt besonders starke Strahlungsunterschiede, zeitlich: zwischen Tag und Nacht, räumlich: zwischen Nord- und Südexposition. Ein Beispiel: Als Extremwerte der Blatt-Temperatur von Ranunculus glacialis während der Vegetationsperiode maß MOSER an einem Grat-Standort 42,1 °C bzw. —8,8 °C.
- Das Temperaturmittel liegt niedriger; die Vegetationsperiode ist kürzer.
- Die Lichtintensität kann bei unverhüllter Sonne und zugleich reflektierenden Wolken die Solarkonstante (1,36 kW/m2), welcher 143 kLux entsprechen, übersteigen.
- Der Anteil an Blaulicht und an UV-Strahlung (UV-A von 400-315 nm und UV-B von 315-280 nm) ist bedeutend höher.
- Die Niederschläge sind höher; sie fallen häufiger als Schnee.
- Die Stürme haben eine starke Umverteilung des Schnees zur Folge: Aperstellen an „Windecken", Graten, Kuppen – Anreicherung in Mulden, Bildung von Wächten in Lee von Rücken.
Die Individuen der Pflanzen müssen also starken zeitlichen Wechsel der Lebensbedingungen bewältigen.
Jan JANSEN schreibt zu den Geröllhalden der Serra da Estrela (2002, 2011):
Ausgehend von den Eiszeiten haben physikalische Verwitterungsprozesse, besonders Frost, an steilen Karwänden und Felsklippen zahlreiche Felspartikel abgesplittert. Einfrieren und Auftauen von Schmelzwasser in Rissen führte zur Dislokation von Gesteinsfragmenten verschiedener Größen. Ein großer Teil des Gesteinsmaterials sammelte sich unterhalb der Klippen an. Die Bereiche mit den angesammelten Gesteinsframenten werden als Schutthalden (engl. screes) bezeichnet. In den Estrela-Schutthalden finden sich auch viele andere Ablagerungen. Aufgrund der Schwerkraft werden die Partikel meist entlang des Hanges sortiert, zumindest dort wo eine große Auswahl an Partikelgrößen zur Verfügung steht. Die größeren Felsen (Felsblöcke) sinken auf die Basis des Hanges, während die winzigen Körner an der Spitze liegen. Unter den heutigen Bedingungen ist in der Serra da Estrela Gesteinsmetereorisation durch Frost von untergeordneter Bedeutung und die meisten der Schutthalden sind eindeutig inaktiv, obwohl es noch einige lokale Aktivitäten aufgrund von physikalischen Prozessen wie Regenabfluss, Spritzwasser und Frostbrüche gibt. Darüber hinaus können aber durch Mensch oder Tiere verursachte Störungen die Schotteroberfläche beeinflussen und Ströme im Schottergestein in Gang setzen. Gute Beispiele von Schutthalden finden sich in der Nähe des Planalto Superior an den Gletschertrögen der Oberläufe des Zêzere und des Alforfa-Flusses.
“Im Untersuchungsgebiet bildet Granit das Hauptgestein oberhalb von etwa 1600 m. Der größte Teil besteht aus eine Verebnungsfläche, die zumeist allmählich bis zum höchsten Kuppenformigen Gipfel (Torre, 1993 m) emporsteigt. Dort aber, wo einige Täler tief in der Plateauverebnung eingeschnitten sind, finden sich einige hochragende Felsen und in ihrer Nähe manchmal eben auch Schutthalden. Die Steinzufuhr ist heute zwar unbedeutend, in manchen Fällen jedoch nicht völlig auf Null reduziert: Frostsprengung, Bildung von Kammeis, Frostmusterböden, usw. werden immer noch beobachtet (Brosche 1971, 1978, Daveau 1973, Daveau et al. 1997). Die Steinzufuhr soll aber während der Kälteeinbrüche im Quartär viel bedeutender gewesen sein und die Schutt-Vegetation hatte in jener Zeit wahrscheinlich eine viel größere Ausdehnung als heute. Von einigen Schuttpflanzen wird angenommen, daß sie seit dem Tertiär hier vorkommen (Braun-Blanquet et al. 1952, Rivas-Martínez 1963).
Eine würmeiszeitliche Vergletscherung ist für die Serra da Estrela nachgewiesen worden (Cabral 1884, Lautensach 1929, Daveau 1971). Die zentrale Hochebene wurde von einem Plateaugletscher norwegischen Typs bedeckt. Es gab sieben Gletscherzungen (Verbreitungskarten in Lautensach 1929, Daveau 1971, Daveau et al. 1997). Die wichtigsten Gletscher waren die der Zêzere, Alforfa und Loriga. Der Zêzeregletscher war der grösste. Er hatte eine Länge von zirka 13 km, reichte bis auf 680 m ü NN in der Nähe von S. Gabriel und dürfte bis 300 m mächtig gewesen sein (Foto 1). Abgesehen von den Gletscherzungen soll die ausdauernde Schneegrenze sich etwa bei 1650 m NN befunden haben. Es ist denkbar, daß damals auch eine typische Vegetation des Gletschervorfeldes existierte. Die Gletscherablagerungen und Moränen sind heute jedoch längst von anderen Vegetationstypen besiedelt.
Die heutige Vegetation der Schutthalden findet ist größtenteils in Karen (portugiesisch Singular: Covão), die von den Gletschern hinterlassen worden sind, besonders in der Nähe der Karwände. Die wichtigsten Fundorte gibt es in den höheren Bereichen (oberhalb 1650 m NN) der Täler von Zêzere, Candieira, Alforfa (Foto 2) und Loriga. Nach Lautensach (1929) ist das Eis, das vom Torreplateau herabkam, wahrscheinlich über die steilen Felswände des oberen Alforfatales herabgebrochen und hat zwischen den Felswänden steile Eisrinnen gebildet. Wahrscheinlich haben ähnliche Ereignisse die Rinnen in den Felswänden des oberen Zêzeretales (Covão do Cimeiro) bedingt. Gerade in diesen Rinnen befinden sich die meisten Schuttgesellschaften. Lautensach (1940, 1942) berechnete, daß die Januartemperaturen Portugals in der Würmeiszeit um fast 6 °C, die Augusttemperaturen um fast 12 °C niedriger lagen als heute. Die Würmeiszeitlichen Niederschlagsmengen sollen dagegen bedeutend höher gewesen sein (Lautensach l.c.).
Die Aufnahmen der Schuttvegetation wurden alle in einem Gebiet gemacht, dessen durchschnittliche Jahresniederschläge heute mehr als 2500 mm betragen (Daveau et al. 1977). Ein Teil davon fällt als Schnee. Auf der Wetterstation Penhas Douradas, die sich auf einer Höhe von 1383 m NN nur 3 km nördlich des Untersuchungsgebietes befindet, wurden während der Periode 1931-1960 jährlich durchschnittlich 34 Tage mit Schneefall und 44 Tage mit Schneedecke beobachtet. Diese Zahlen sind in den Karen auf der Ostseite des noch 600 m höheren Plateaus wohl viel höher. So hat es am Gipfel sogar noch am 29. Juni 1997 geschneit. In günstigen Lagen kann man Schneeflecken bis in den frühen Sommer hinein beobachten. Der Herbst ist im allgemeinen die niederschlagsreichste Jahreszeit, jedoch sind die Niederschläge auch im Winter und im Frühjahr bedeutend. In den Monaten Juli und August sind sie niedrig und zeigen damit den mediterranen Einfluß an.
Dieser äußert sich ebenfalls in den hohen mittleren Jahressummen der Sonnenscheindauer. Auf Penhas Douradas werden mehr als 2500 Stunden Sonnenschein pro Jahr registriert. Die Jahresmitteltemperatur an der Wetterstation beträgt 8,9 °C, mit einem mittleren täglichen Minimum von -0,4 °C im Januar und einem mittleren Tagesmittel von 17,2 °C im Juli (Amorin Ferreira 1965). Die Minimumtemperatur taucht dort durchschnittlich an 66 Tagen im Jahr unter den Gefrierpunkt; auf 1510 m Höhe in Penhas da Saúde, also etwa 100 m unterhalb des Untersuchungsgebietes, ergeben sich schon 82 Tage im Jahr (Amorin Ferreira 1965). Auswirkungen des Klimas zeigen im Untersuchungsgebiet eine asymmetrische Verteilung, in bezug auf die Schneefälle z.B. eine West-Ost Asymmetrie (Lautensach 1929, Daveau 1971, 1986). Die Anhäufung der Schneemassen an der Ostseite des Gebirges lässt sich durch die vorherrschende Westwinde erklären. Außerdem gibt es eine Nord-Süd Asymmetrie: Wegen der stärkerer Einstrahlung der Sonne sind die süd-exponierten Gletscherzungen kürzer (Daveau 1971). Die Verbreitung der Schuttgesellschaften läßt sich analog erklären. Der größte Teil befindet sich nämlich in den ost-exponierten oberen Bereichen der ehemaligen Gletscher der Zêzere, Alforfa und Candieira.” (Jan JANSEN, 2011)
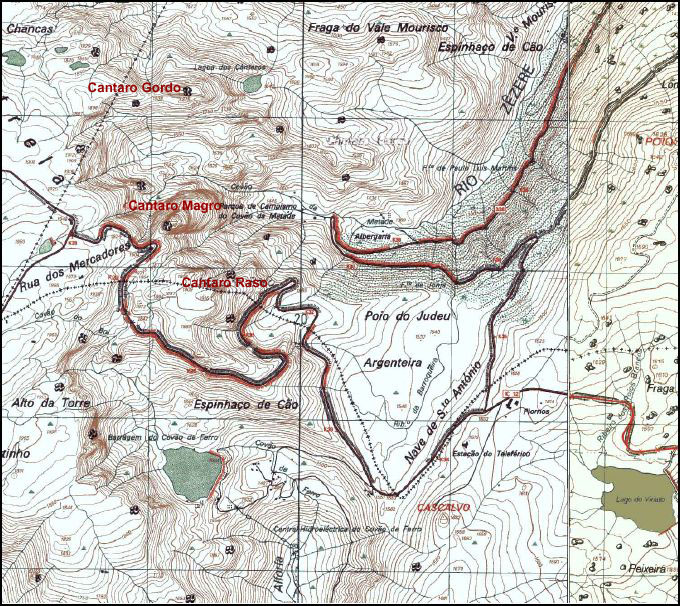
Hochebene (Planalto) der Serra da Estrela mit den 3 Cântaros (Felsspitzen) und Oberläufe (Gletschertröge) von Zêzere und Alforfa (Kartenausschnitt: Cartas Topográficas de Portugal. 1:25.000)
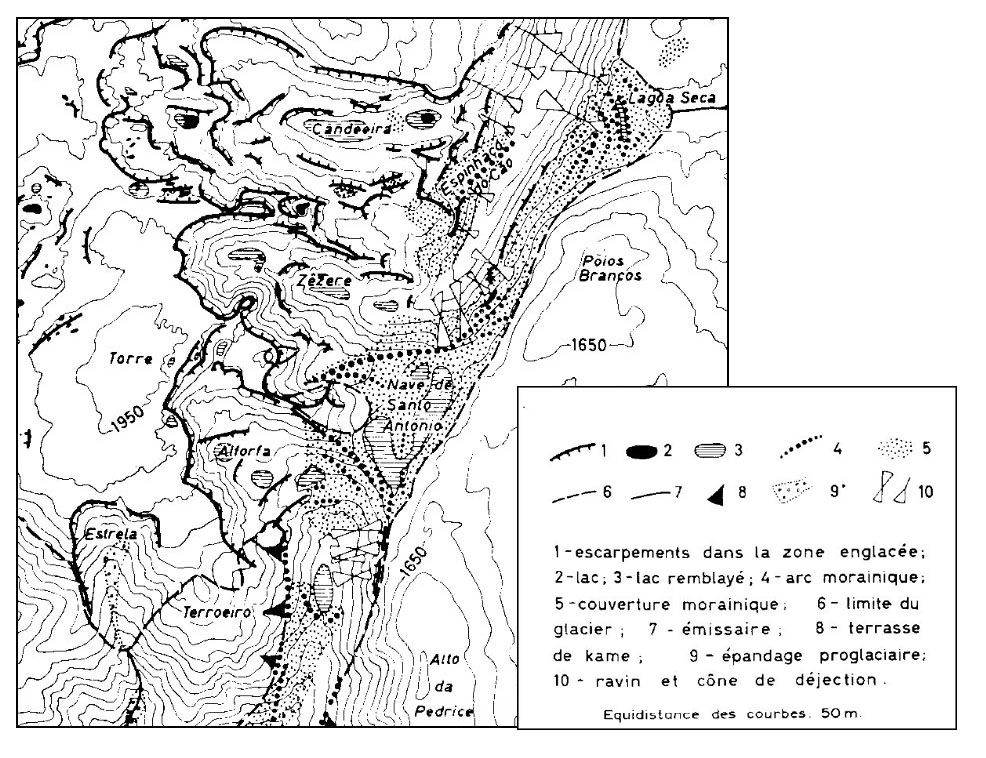
Würm-Eiszeitliche Vergletscherung der Serra da Estrela (Nach: DAVEAU, S. (1971) )
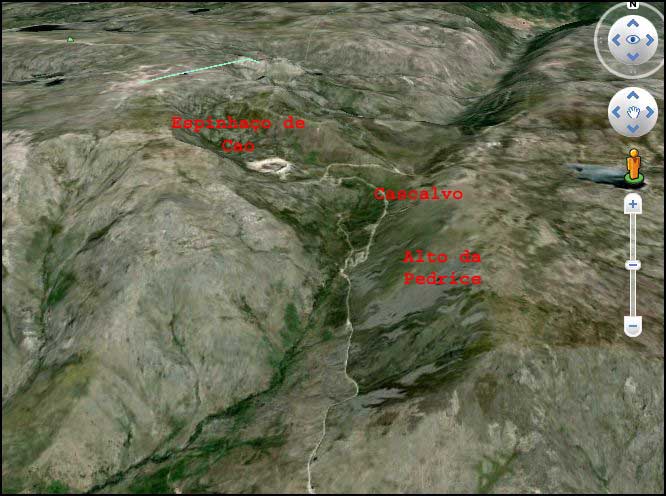
Die Gebiete “Espinhaço do Cão” und “Cascalvo” der Ribeiro do Alforfa, wo es mediterrane orophile Silikatfels-Schutthalden gibt. “Alto da Pedrice”, ein enormes Blockhaldenfeld im Trogtal des Alforfa, is auch im Satellitenfoto (Google-Earth) deutlich erkennbar.

Blick auf das Tal des Alforfa-Flusses (Serra da Estrela). Am Grunde die Blockhalden des Alto da Pedrice. In der Mitte des Fotos das Wasserrückhaltebecken: Barragem do Covão de Ferro.

Cântaro Gordo - Südflanke des Cântaro Gordo (Serra da Estrela) mit Trogtal (Kar) des Zêzere-Flusses (der Pfeil verweist auf eine Schutthalde (engl. scree)).

Lagoa dos Cântaros (Links) und Nordflanke des Cântaro Gordo (Mitte) mit Schutthalden (grau im Foto) (Satellitenfoto: Google Earth)
|

Links im Bild die Schutthalden der Nordflanke des Cântaro Gordo - (An der Nordflanke des Cântaro Gorde findet sich die seltene Assoziation des Cryptogrammo-Dryopteridetum mit Cryptogramma crispa.)
|
 |
 |
 |

Covão do Cântaro e Lagoa dos Cântaros (Serra da Estrela)
|
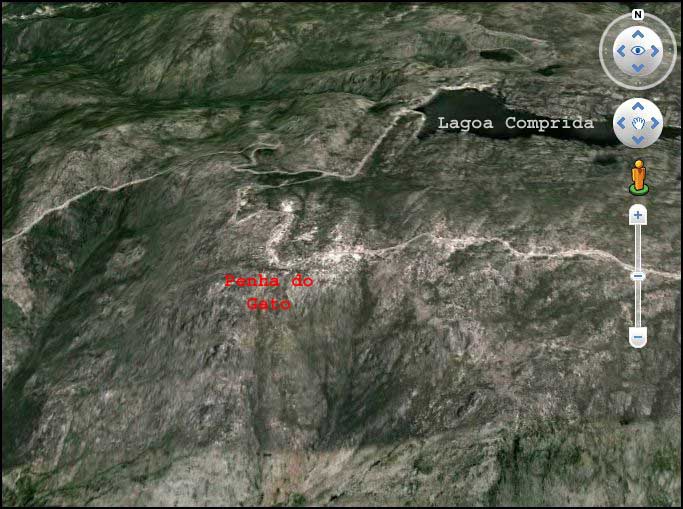
Gebiet der “Penha do Gato” oberhalb der Lagoa Comprida, wo es ebenfalls inaktive mediterrane orophile Schutthalden gibt (Satellitenfoto: Google-Earth)

Die meisten der Schutthalden sind mäßig bis stark geneigt, bei überwiegend nördlicher bis östlicher Exposition. Sie liegen oft zwischen steilen Felswänden, die weitgehend das Eindringen von Sonnenschein verhindern. Daher bleibt die Schneedecke länger erhalten. Die Topographie begünstigt eine Schneeansammlung, da starke Winde den Schnee über die Hochebene auf die Leeseite der Felskliffe (wie z.B. die Nordflanke des Cântaro Gordo) blasen, wo die Schutthalden auftreten. Die Schneebedeckung gibt Schutz vor Kälte, vor frühem Auskeimen der Pflanzen und vor Austrocknung. Darüber hinaus erhöhen Schneedecke und Schneeschmelze die Bodenfeuchtigkeit. Der Klimawandel (insbesondere Veränderungen in Schneefallhäufigkeit und Frostwirkung) kann diese Biotope wesentlich beeinflussen. (Die Lebensbedingungen in den Schutthalden sind für Pflanzen extrem. Das Bewegen einzelner Gesteinspartikel oder das Gleiten des ganzen Hanges kann die Wurzelsysteme von Pflanzen beschädigen oder sie begraben. Die Quasi-Abwesenheit von feiner Erde in der oberen Hangschicht gibt Samen nur eine sehr geringe Möglichkeit, zu keimen. Verschiedene Wachstumsformen treten auf, wobei aber im allgemeinen Schutthaldenspezialisten lange, vertikale und/oder horizontale Wurzeln besitzen, um mit dem abrutschenden Material fertig werden zu können. Ein kreisförmiges Muster von feinen Wurzeln ermöglicht es ihnen, Wasser und Nährstoffe aufzunehmen. Die meisten Pflanzen besitzen Stolonen und kriechende Rhizome. Nach SCHRÖTER (1926) können fünf grundlegende Wachstumsformen unterschieden werden: 1. Schuttwanderer, 2. Schuttüberkriecher, 3. Schuttstrecker, 4. Schuttdecker, 5. Schuttstauer. Einige Pflanzen können Zwischenmerkmale haben.
Sobald einige Pioniere sich niedergelassen haben, können sie organisches Material und Humus ansammeln. Wenn nicht erneut viele zusätzliche Felstrümmer niederstürzen, kann weitere Vegetation Fuss fassen und sich eventuell zu Grasland oder Gebüsch entwickeln. Im allgemeinen nehmen diese Prozesse Tausende von Jahren in Anspruch. Große Blockhalden werden kaum von Pflanzen kolonisiert, weil es kein feines Material für die Wurzelbildung gibt. Ein gutes Beispiel ist die riesige Blockhalde am westlichen Hang des Alto da Pedrice (siehe das Foto oben)
Einige Schutthaldenspezialisten treten in sekundären Biotopen mit hoher Störung auf, wie z.B. verbrannte Gebiete, Straßenböschungen, künstliche Lichtungen und Steinbrüche. In den höheren Zonen der Serra da Estrela können mindestens zwanzig für Schutthalden charakteristische Pflanzenarten (sog. "Scree-Spezialisten") gefunden werden. Neun von ihnen (markiert mit *) treten auf der vorläufigen roten Liste der Gefäßpflanzen in Portugal auf; dreizehn (+) sind in Portugal auf die Serra da Estrela beschränkt; eine (▲) ist eine strikt endemische Spezies der Serra da Estrela. Viele dieser Pflanzenarten könnten aus dem Tertiär stammen.
Folgende Arten sind charakteristisch für die Schutthalden der Estrela:
(Charakterarten von Syntaxa der Klasse Thlaspietea rotundundifolii (Rivas-Martínez 1981b, RivasMartínez et al. 1984, 1989) - mit (*) markierte Arten treten auf der vorläufigen roten Liste der Gefäßpflanzen in Portugal auf; mit (+) markiere Arten sind in Portugal auf die Serra da Estrela beschränkt; eine (▲) Art ist eine strikt endemische Spezies der Serra da Estrela)
 | |
 |  |
 |  |
 |  |
 |  |
 |  |
 | 
Reseda gredensis * +
|
 | 
Scrophularia herminii
|
 |  |
 |   |
 |  |
.
Von den obengenannten, insgesamt 20 Arten stehen 9 (mit ● markiert) auf der vorläufigen Roten Liste der Gefäßpflanzen Portugals (Ramos Lopes et al. 1990); 13 (+) sind innerhalb Portugals auf die Serra da Estrela beschränkt, unter denen ein Endemit (▲). Es handelt sich meistens um biotoptypische stenotope Arten die, weil die Biotoptypen nur in geringer Gesamtfläche bzw Anzahl vorkommen, potentiell gefährdet sind. Die meisten Arten gehören wohl zu isolierten Restpopulationen, die früher wahrscheinlich eine größere Ausdehnung hatten. Einige Arten, vor allem Arrhenatherum cf. carpetanum, Coincya orophila, Lactea viminea und Linaria saxatilis konnten auch Sekundärstandorte besiedeln, wodurch ihr Areal erweitert wurde.
Doronicum carpetanum soll in der Serra da Estrela vorkommen (Franco 1984). Jan JANSEN (2011) konnte diese Art jedoch nicht mehr nachweisen, und nimmt an, dass sie in der Estrela ausgestorben ist. Von der nahe verwandten Art die, rezent von Pérez-Morales et al. (1994) als Doronicum pubescens beschrieben wurde, konnte Jan JANSEN jedoch 1996 sieben Exemplare an einer Stelle am Wegrand im Genistion polygaliphyllae unweit von relativ frischen Betula alba-Beständen auf einer Höhe von 1400 m beobachten (Aufnahme 96-109).
Pflanzensoziologische Übersicht über die Schutthaldenvegetation der Serra da Estrela
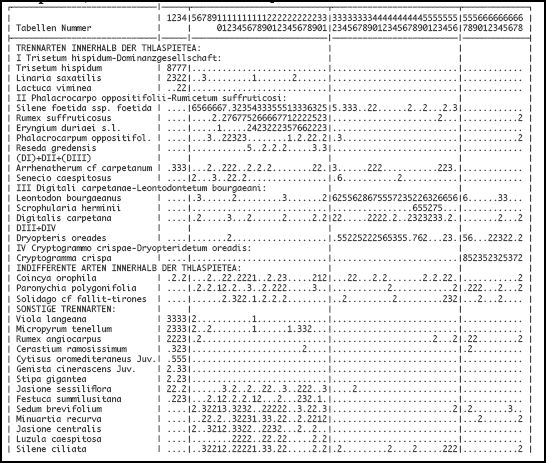
Jan JANSEN (1998, 2011) kann aufgrund pflanzensoziologischer Erhebungen (siehe Annex 1) folgende vier Gemeinschaften innerhalb der Schutthaldenvegetation (Klasse Thlaspietea rotundundifolii) der Serra da Estrela unterscheiden:
- Trisetum hispidum-Dominanzgesellschaft
- Sileno foetidae-Rumicetum suffruticosi (Rivas-Martínez) Jansen ass. nov.
- Digitali carpetanae-Leontodontetum bourgaeani ass. nov. hoc loco
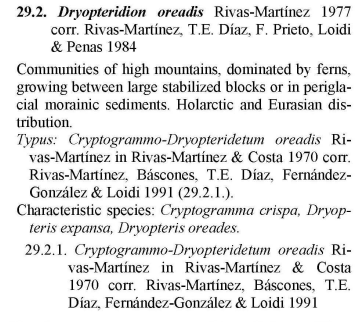
- Cryptogrammo crispae-Dryopteridetum oreadis Rivas-Martínez in Rivas-Martínez & Costa 1970
Außerdem beschreibt er noch eine weitere Schutthaldengesellschaft, das
- Violetum langeanae Ortiz & Marcos Samaniego 1989
für die Serra da Estrela, verzichtet aber wegen noch unzureichender Kenntnisse über diese Assoziation auf eine genauere phytosoziologische Zuordnung.
2.1 Trisetum hispidum Dominanzgesellschaft (siehe Tabelle 1 in Annex 1)
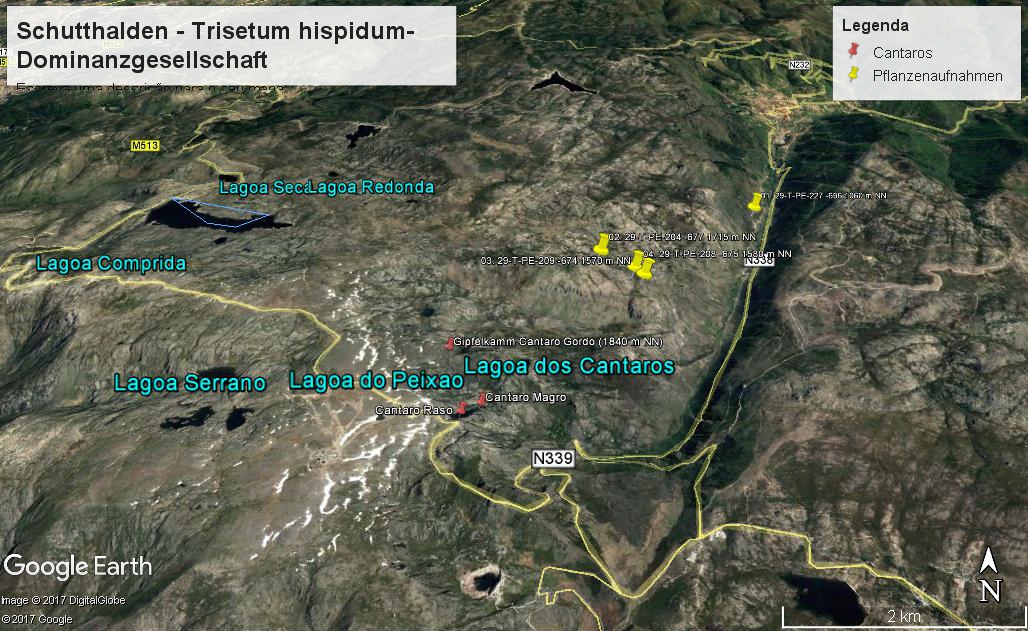
Vorkommen der Trisetum hispidum Dominanzgesellschaft in der Serra der Estrela (Vegetationsaufnahmen: Jan JANSEN 2011)
Diese Gesellschaft wird durch die Dominanz von Trisetum hispidum (Trisetaria hispida) sowie durch das regelmäßige Auftreten von Linaria saxatilis charakterisiert. Lactuca viminea und Arrhenatherum cf. carpetanum, zwei Thlaspietea-Arten, differenzieren die Gesellschaft gegenüber den anderen dieser Klasse. Einschließlich Coincya orophila gibt es in allen vier Aufnahmen nur fünf Thlaspietea-Arten. Wie im Violetum langeanae treten auch in der Trisetum hispidum-Dominanzgesellschaft viele Arten auf, die in Gesellschaften der Koelerio-Corynephoretea und Tuberarietea verbreitet sind. Nicht die Zahl sondern die Gesamtdeckung der Thlaspietea-Arten ist größer als die der Koelerio-Corynephoretea- und Tuberarietea-Arten. Dies wird insbesondere durch die Dominanz von Trisetum hispidum verursacht. (Jan JANSEN 2011)
Charakteristische Arten:
| |
 |  |
 |  |
 |  |
2.2 Silene foetida - Schutthaldenvegetation (siehe Tabelle 1 in Annex 1)
(Sileno foetidae-Rumicetum suffruticosi (Rivas-Martínez) Jansen ass. nov.)
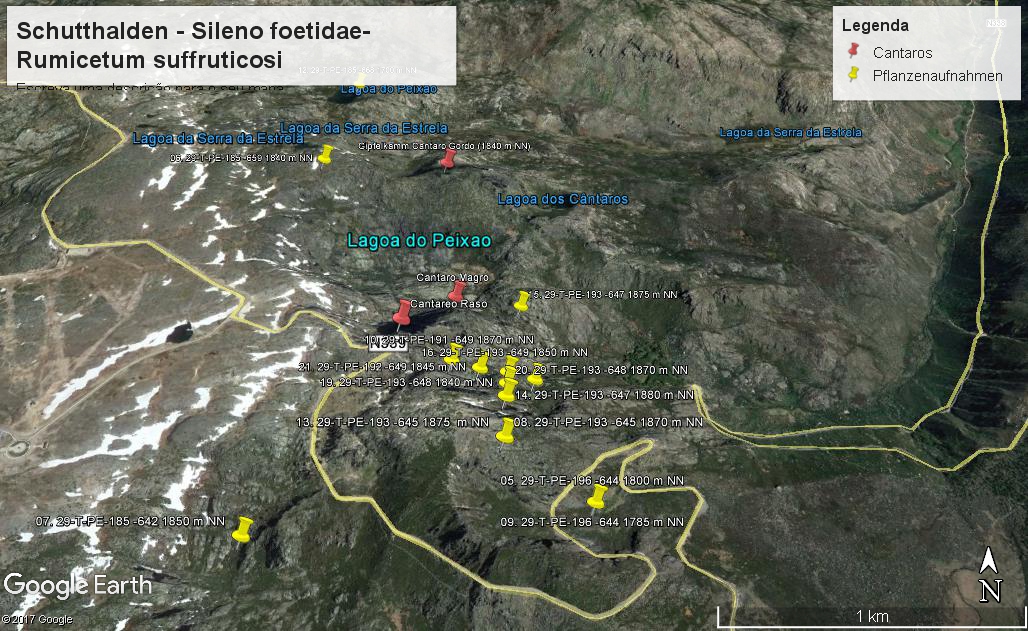
Vorkommen der Silene foetida - Schutthaldenvegetation in der Serra der Estrela (Vegetationsaufnahmen: Jan JANSEN 2011)
Das Sileno foetidae-Rumicetum suffruticosi ist eine meso- bis xerophytische Dauergesellschaft auf granitischen Schutthalden. Die relativ offene Gesellschaft ist beschränkt auf das Estrela-Gebirge, wo sie oberhalb 1600 m NN nahe an den Karwanden der ehemaligen Gletschertäler der Zêzere und Alforfa vorkommt. Die relativ seltenen Standorte sind oft bis in den Frühling hinein mit Schnee bedeckt. Die Gesellschaft zeigt ihr phänologisches Optimum erst im Vollsommer (abhängig von den Wetterbedingungen ungefähr Ende Juli bis Ende August). Die Schutthalden sind überwiegend aus einer Mischung von Fein- und Grobschutt aufgebaut, obwohl Blöcke fast nie fehlen
Silene foetida-Schutthalden sind relativ trockene Schutthalden, bestehend aus kleinen bis mittelgroßen, teilweise verfestigten Gesteinen. Silene foetida subsp. foetida ist ein streng endemisches Taxon der Serra da Estrela. Silene foetida subsp. gayana ist ein verwandtes Taxon endemisch für NW-Spanien, welches ebenfalls Schutthalden kolonisiert. Silene foetida subsp. foetida durchspinnt den Schutt mit langen Trieben, die dann wieder wurzeln (Schuttwanderer). Auf diese Weise bewegt sie sich entlang der gleitenden Felsmaterialien. Die Pflanze ist klebrig und erhöht so ihre Fähigkeit, sich an den Trümmern zu halten. Andere Arten, die für diese Umgebung typisch sind, sind Eryngium duriaei subsp. duriaei, Phalacrocarpum oppositifolium, Reseda gredensis und Rumex suffruticosus. Alle von ihnen sind auf den oberen Gürtel mit Ausnahme von Phalacrocarpum oppositifolium beschränkt. Diese hübsche Komposite wächst auch in Felsspalten und Talushängen des Mittleren Gürtels. Ihre verbreitung umfasst mehr oder weniger den nordwestlichen Quadranten der Iberischen Halbinsel. Rumex suffruticosus ist eine iberischer Endemismus, der oft mit Trisetaria hispida in den Gebirgen von Nordwest-Spanien wächst. Das ist bei der Estrela nicht der Fall. Hier scheint letztere die Dynamik der Brandgebiete zu bevorzugen, wobei sie manchmal die post-fire-Vegetation an Hängen des oberen Abschnittes des Zêzere-Tals dominiert.
Die Wachstumsstrategie von Eryngium duriaei subsp. duriaei unterscheidet sich von der von Silene foetida. Die Pflanze besitzt einen langen und dicken Wurzelstock, der als Anker tief in den Schutt wächst und die bewegten Materialien hochhält ("Talustrage"), Reseda gredensis ist äußerst selten und wird durch eine geschätzte Population von weniger als hundert Exemplaren repräsentiert. Es ist eine endemische Art, die nur aus der Estrela und der Gredos bekannt ist. Silene foetida Schutthalden können in offenes xerophytisches Gipfel-Grasland in Bereichen, wo es eine größere Anhäufung von Kies und anderem feinen Material gibt, übergehen.
Am Rande der Schutthalden, meistens an nord-exponierten und relativ feinerdereichen Stellen, grenzt das Sileno foetidae-Rumicetum suffruticosi manchmal an eine, wenigstens in der Serra da Estrela noch nicht beschriebene Gesellschaft, die meistens von Luzula caespitosa oder Silene ciliata dominiert wird und zu den Festucetalia indigestae gehört. Auch das stete Auftreten von Minuartia recurva und die relativ hohe Präsenz von Jasione centralis zeigen diese Verbindung mit den psychro-xerophytischen Trockenrasen.
Direkt unterhalb hoher Felsen, ebenfalls auf nord-exponierten Stellen, wo der Schnee am längsten liegen bleibt und auch im Hochsommer die Vegetation der Sonne nur relativ kurz ausgesetzt ist, grenzt unsere Gesellschaft öfter an eine chionophytische Nardetea-Gesellschaft, die nur unvollständig als Campanulo-Festucetum beschrieben worden ist (Rivas-Martínez 1981b, Jansen 1997). Zu den anderen Arten die relativ gut an sommertrockenen Verhältnissen adaptiert sind, zählen Jasione sessiliflora, Micropyrum tenellum, Arenaria querioides, Sedum brevifolium.
Die Felswände tragen eine Vegetation die meistens zum Murbeckiello-Saxifragetum (Asplenietea) gehört. An sonnig exponierten Felsen gibt es Ansätze zum sogenannten Sileno foetidae-Dianthetum lusitani. Bis jetzt habe ich nur Aufnahmen der Felsvegetation der Hoch-Estrela gesammelt. Eine Beschreibung steht noch aus, aber aus meinen Geländebeobachtungen habe ich feststellen können, daß Silene foetida überhaupt nicht im Sileno-Dianthetum vorkommt. Statt dessen tritt öfter Silene acutifolia auf. Es dürfte sich dabei um eine Verwechslung dieser beiden Arten handeln.
In den niedrigen Bereichen grenzt die Halde an Bestände, in denen vor allem Erica arborea dominiert. Diese Bestände gehören meistens zur Klasse Pino-Juniperetea.
An relativ feuchten Stellen (Umgebung von Brunnen, zeitweise wasserführende Bäche, große Blöcke) leitet die Gesellschaft zum Farn- und oft Hochstauden-reichem Digitali-Leontodontetum über.
Das Sileno foetidae-Rumicetum suffruticosi ist eine Dauergesellschaft. Nur am Rande der Schutthalden kann es Entwicklungen geben, die zu einigen der oben genannten Gesellschaften führen.
Die Gesellschaft ist wegen ihrer geringen Gesamtfläche potentiell gefährdet. Sie wird unregelmäßig von Schafen, Ziegen und letzter Zeit vor allem von Rindern besucht. In wie fern sie hierdurch oder durch Klimaänderung (vor allem Änderungen in Schneefall und Frostaktivität) beeinträchtigt wird, ist nicht bekannt.
Folgende Arten sind charakteristisch für diese Gesellschaft:
 |  |
 |  |
 |
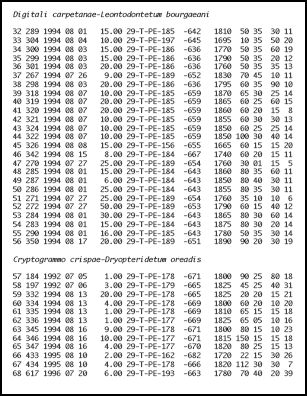
2.3 Leontodon bourgaeanus - Schutthaldenvegetation (siehe Tabelle 1 in Annex 1)
(Digitali carpetanae-Leontodontetum bourgaeani ass. nov. hoc loco)

Vorkommen der Leontodon bourgaeanus - Schutthaldenvegetation in der Serra der Estrela (Vegetationsaufnahmen: Jan JANSEN 2011)
Die Gesellschaft wird durch das stete Auftreten von Leontodon bourgaeanus und Dryopteris oreades charakterisiert. Bezeichnend ist auch das häufige Auftreten von Digitalis carpetana und die Präsenz der relativ seltenen Scrophularia herminii. Angelica major ist eine gute Differentialart zu den anderen in Tab. 1 dargestellten Einheiten. Sie tritt aber auch regelmäßig in Gesellschaften der Montio-Cardaminetea und Molinio-Arrhenatheretea auf.
Vegetation mit Leontodon hispidus subsp. bourgaeanus besiedelt Schutthalden, die zusätzlich mit aufgelösten Nährstoffen von den Weiden der Hochebene (planalto superior) versorgt werden. Diese Schutthalden können zum Teil als zeitlich begrenzte Bachbetten fungieren, die Schmelz- und Regenwasser transportieren.
Das Digitali-Leontodontetum ist eine meso- bis hygrophytische Dauergesellschaft, die auf granitischen Schutthalden wächst. Die relativ offene Gesellschaft ist auf das Estrela-Gebirge beschränkt, wo sie oberhalb 1650 m NN nahe an den Karwänden der ehemaligen Gletschertäler der Zêzere, Alforfa und Loriga vorkommt. Die relativ seltenen Standorte sind oft bis in den Frühling hinein mit Schnee bedeckt. Die Standorte der Gesellschaft apern relativ spät aus, und die Gesellschaft zeigt ihr phänologisches Optimum erst im Vollsommer (abhängig von den Wetterbedingungen ungefähr Ende Juli bis Ende August). Die relativ feinerdearmen bis mäßig feinerdereichen Schutthalden sind überwiegend aus einer Mischung von Grob- und Blockschutt aufgebaut.
Einzelne Pflanzen von Leontodon bourgaeanus sind häufig miteinander verwoben und bilden kleine Matten ('talus coverer'). Diese werden oft von Digitalis purpurea subsp. carpetana und dem Wurmfarn Dryopteris oreades begleitet. In solch einer feuchten Umgebung können große Hochstauden wie Scrophularia herminii, Angelica major und Crepis lampsanoides gedeihen, besonders an Gebieten, die vor starken Winden und langen Sonnenzeiten geschützt sind. Doronicum carpetanum muss auch in diesem Biotop gewachsen sein. Es scheint jedoch, dass diese große Hochstauden-Spezies inzwischen ausgestorben ist. Die verwandten Doronicum pubescens und Doronicum plantagineum treten nicht im gleichen Biotop auf. Erstere wurde vor kurzem beschrieben und neu für die Flora der Serra de Montesinho Nordwest-Spaniens beschrieben. Bisher wurde sie in der Estrela nur einmal gefunden, in Genista florida Gebüsch im Mondego-Becken gelegen. Unter ähnlichen Wachstumsbedingungen könnte man die Art aber auch in Birkenwäldern erwarten. Doronicum plantagineum kann in Ständen der Pyrenäen-Eiche (Quercus pyrenaica) entlang der Straße nach Poço do Inferno beobachtet werden.
In weniger dynamischen Gebieten, oft in der Nähe von Felsnasen oder an sanft geneigten Standorten kann Leontodon bourgaeanus-Geröllvegetation in Festuca henriquesii-Wiesen übergehen.
Charakteristische Arten:
| |
 |  |
 | 
Scrophularia herminii
|
2.4 Cryptogramma crispa - Schutthaldenvegetation (siehe Tabelle 1 in Annex 1)
(Cryptogrammo crispae-Dryopteridetum oreadis Rivas-Martínez in Rivas-Martínez & Costa 1970)
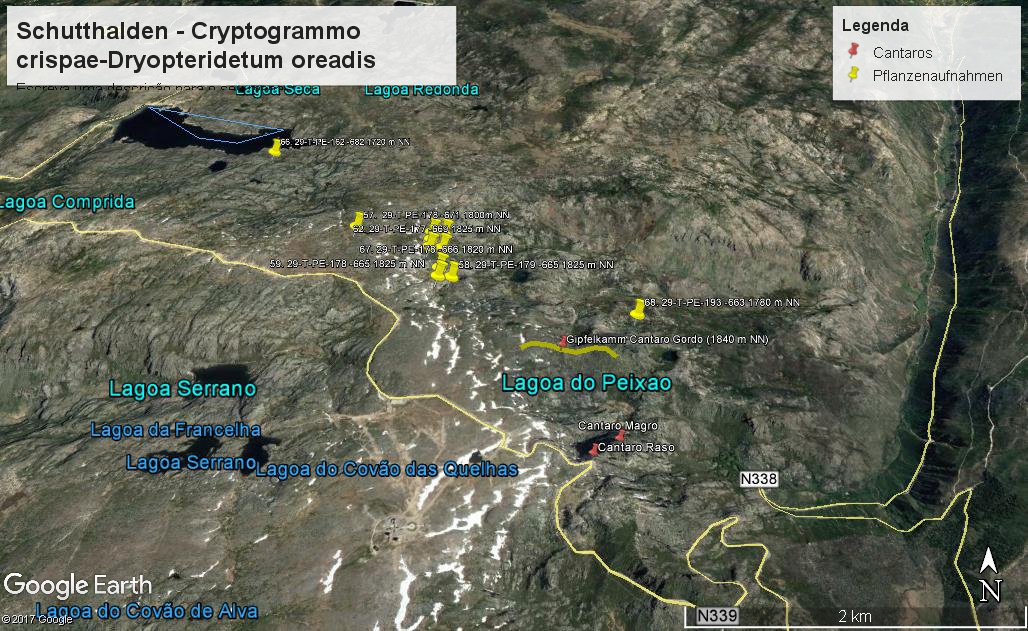
Vorkommen der Cryptogramma crispa - Schutthaldenvegetation in der Serra der Estrela. (Vegetationsaufnahmen: Jan JANSEN 2011)
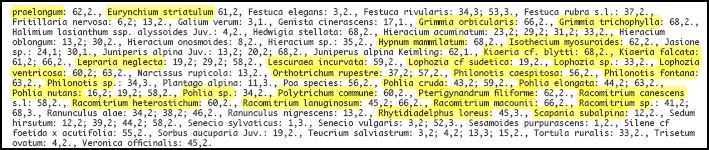
Die Gesellschaft wird in der Serra da Estrela durch den Krausen Rollfarn Cryptogramma crispa charakterisiert. Folgende Arten differenzieren gegen die übrigen Schuttgesellschaften: Sedum anglicum, Nardus stricta, Silene acutifolia, Galium saxatile, und viele Moose wie Polytrichastrum alpinum, Kiaeria starkei, Pseudotaxiphyllum elegans, Dryptodon patens, Plagiothecium denticulatum und Barbilophozia floerkei. Die Affinität mit dem Digitali-Leontodontetum drückt sich im Vorkommen folgender gemeinsamer Arten aus: Dryopteris oreades, Agrostis castellana und die Moose Lescuraea patens und Brachythecium dieckii.
Meistens bleiben Moose und Flechten in Aufnahmen auf der Iberischen Halbinsel unberücksichtigt. Deshalb ist es schwierig, ihre pflanzensoziologische Stellung anzugeben. Selbst in der Estrela ist es zur Zeit nicht möglich, lokale Charakterarten zu unterscheiden, weil bisher eine syntaxonomische Übersicht fehlt. Bis auf weiteres kann bei den Moosen nur von lokalen Differentialarten die Rede sein.
Diese Art von Vegetation ist sehr selten, bewohnt kleine Standorte in mehr oder weniger konsolidierten Schutthalden, bestehend aus mittelgroßen bis großen Felsen. Der Krause Rollfarn (Cryptogramma crispa) ist ein Gletscherrelikt aus Europa und Westasien. In der Estrela bevorzugt er schattige Bedingungen, die meistens in der Nähe großer Felsbrocken gefunden wurden. Es wird davon ausgegangen, dass die Spezies weitgehend von dem Schutz abhängt, den die Schneedecke gegen Frost bietet.
Jan JANSEN schreibt zum Vorkommen dieser Gesellschaft in der Estrela:
Ich habe das Cryptogrammo-Dryopteridetum fast nur in Karen nördlich des Cântaro Gordos gefunden, wo einst der Candieiragletscher wurzelte. Diese Stellen sind im Winter relativ gut gegen Kälte geschützt, weil sie dann oft von Schnee bedeckt sind. Festuca henriquesii (eigene Beobachtungen) und Kiaeria starkei (Frey et al. 1995) können als Schneezeiger gelten. Auch diese Gesellschaft zeigt ihr phänologisches Optimum auf Grund der langen Schneebedeckung erst im Spätsommer (August). (Jan Jansen, 2001)
Vegetation mit Krausem Rollfarn beherbergt interessante Bryophyten, von denen einige auch Gletscher-Relikte sind. Die häufigste Farn-Spezies in dieser Umgebung ist der auffällig getuftete Geröll Wurmfarn (Dryopteris oreades) und so ist es aus der Ferne schwierig, den Cryptogramma crispa-Vegetationstyp von dem Leontodon Bourgaeanus-Vegetationstyp zu unterscheiden. Nur aus nächster Nähe erkennt man den kleinen Krausen Rollfarn und einige der kleinsten Bryophyten. Cryptogramma crispa-Geröllvegetation ist in der Regel viel besser in den höheren iberischen Gebirgen, die während der vorletzten Kaltzeit vereist waren, entwickelt als in der Estrela. Leontodon bourgaeanus-Vegetation scheint dagegen am besten in der Estrela entwickelt zu sein. Dies ist vermutlich auf die Unregelmäßigkeit sowohl von Schneefall als auch der Schneedecke in der Estrela zurückzuführen. Der Feingliedrige Dornfarn (Dryopteris expansa) ist vielleicht noch seltener als der Krause Rollfarn. In Ausnahmefällen können sie zusammen wachsen. Allerdings wird Dryopteris expansa auch in der Nähe von Saxifraga stellans-Quellen gefunden. In Portugal sind der Krause Rollfarn (Cryptogramma crispa) und der Feingliedrige Dornfarn (Dryopteris expansa) auf den Oberen Gürtel der Serra da Estrela beschränkt (siehe auch RIVAS-MARTINEZ et al. 1989). Dryopteris oreades tritt dagegen auch in den höheren Gebieten der Serra do Gerês auf.
Jan JANSEN schreibt zum Vorkommen von Dryopteris expansa in der Estrela:
Ich habe den Farn nur in oder in der Nähe von Quellen angetroffen. Die Art scheint in der Estrela also nicht auf Schutthalden beschränkt zu sein. Im Cryptogrammo-Dryopteridetum selbst habe ich diese Art nie beobachtet.) (J. Jansen, 2001)
.
Charakteristische Arten:
| |
 |  |
Differentialarten zu den anderen Schutthaldengesellschaften:
| |
 |  |
 |  |
Davon die folgenden Moosarten:
| |
Dryptodon patens
| |
Begleiter von Schutthaldengesellschaften:
| |
 |  |
 |  |

Deschampsia flexuosa (iberica)
|  |
 |  |
 |  |
   |
 |
 |
 |
2.5 Violetum langeanae Ortiz & Marcos Samaniego 1989 (siehe Tabelle 2 in Annex 1)
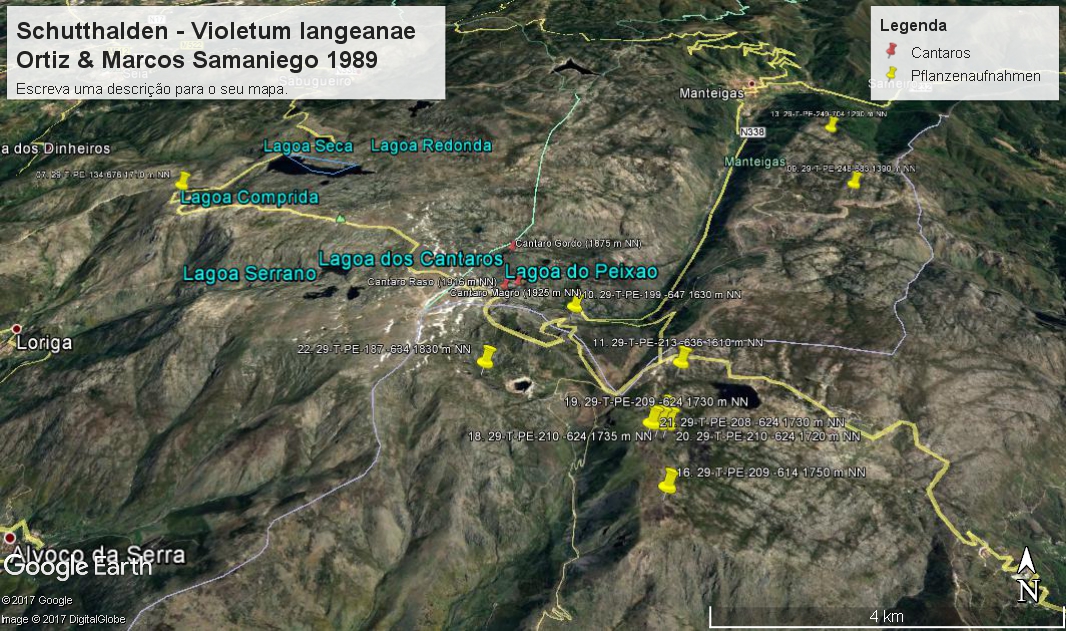
Vorkommen des Violetum langeanae in der Serra der Estrela (Vegetationsaufnahmen: Jan JANSEN 2011)
Das Violetum langeanae wird durch Viola langeana charakterisiert in Verbindung mit Arten, die in lückigen Pionier-Gesellschaften auf offenen Trockenrasen, Sand- und Grusfluren allgemein verbreitet sind: Agrostis truncatula, Arenaria querioides, Jasione sessiliflora, Rumex angiocarpus, Sedum brevifolium, Micropyrum tenellum und viele andere. Einige Arten, die ihr Optimum in Schuttfluren haben, können vereinzelt vorhanden sein, wie Arrhenatherum cf. carpetanum und Linaria saxatilis.
Zwei Varianten lassen sich unterscheiden:
- Variante mit Galium vivianum
In der letztgenannten wird außerdem eine Subvariante mit Cytisus oromediterraneus abgetrennt. Trennarten der Variante mit Galium vivianum sind Galium vivianum, Paronychia polygonifolia und Hypochaeris radicata. Außerdem sind vielleicht auch Sedum brevifolium, Festuca summilusitana und Conopodium majus als Differentialarten innerhalb der Assoziation zu betrachten.
Ortiz und Marcos Samaniego (l.c.) haben das Violetum langeanae den Thlaspietea zugeordnet. Die Anzahl und Deckung vieler Arten, die in der Serra da Estrela und (ähnlichen benachbarten Gebirgen des Sistema Central) auf Grus- und Sandfluren vorkommen, sind aber viel bedeutender.
COSTA et al. 2012 ordnen das Violetum langianae ebenfalls der Klasse Thlaspietea zu:
 |  |
 |  |
Aus Tabelle 2 (Annex 1) (Jan JANSEN 2011) geht jedoch hervor, daß vor allem Arten der Klassen Koelerio-Corynephoretea und Tuberarietea in den Pflanzenaufnahmen, die Viola langeana enthalten, vorherrschen; solche der Klassen Thlaspietea und Ruderali-Secalietea sind weniger vertreten. Eine Zuordnung des Violetum langeana zu der Klasse Thlaspietea erscheint deshalb nach Jan JANSEN (2011) fraglich und bedarf einer Überprüfung.
Das auf die Westabschnitte der Zentralkordillere (Iberisches Scheidegebirge) beschränkte, gelbblühende Stiefmütterchen Viola langeana wächst in der Serra da Estrela wahrscheinlich primär auf granitischen Grusfluren, die durch natürlichen Erosionsprozesse entstanden sind. Der Granitgrus entsteht vor allem durch Spaltenfrost bzw. Temperatursprengung aus dem anstehenden Gestein oder aus großen (manchmal erratischen) Blöcken, die z.T. massenhaft vorhanden sind.
Der im Vegetationsmuster befindliche Grus kann an Ort und Stelle an der Felsoberfläche entstanden oder auch durch Wind und Wasser transportiert worden sein. Im Bereich des Hochplateaus sind die Böschungen im allgemeinen oft flach, sodaß vor allem dort grusige Massen entstehen. Viola langeana wächst auch an sekundären Standorten, wie Straßenböschungen, Steingruben, Äckern und Brandflächen (s. Jan JANSEN 1994b).
Die Variante mit Galium vivianum wächst im allgemeinen auf Rohböden, die zum größten Teil aus fein- bis grobkörnigem Feinschutt aufgebaut sind, dem wenig Sand beigemischt ist. Dies ist möglicherweise der Grund, warum Paronychia polygonifolia die Standorte dieser Variante zu bevorzugen scheint. Nach RIVAS-MARTÍNEZ et al. (1989) ist Paronychia polygonifolia Charakterart der Androsacetalia alpinae, Ordnung der Silikatschuttgesellschaften. Die Variante mit Galium vivianum scheint die oben beschriebenen primären Standorte zu bevorzugen.
Den Böden der Variante mit Corynephorus canescens ist meistens mehr Sand beigemischt. Die Subvariante mit Cytisus oromediterraneus findet sich auf rezenten Brandflächen, wo die Asche von verbranntem organischem Material sich mit mineralischem Sand und Granitteilchen mischt. Hier kann es öfters zur Flächenerosion kommen, bei der die kleinen Partikel zuerst abgetragen werden.
Nach ORTIZ, S. & N. M. SAMANIEGO (1989) sind die Standorte der Variante mit Galium vivianum relativ feucht und die mit Corynephorus canescens relativ trocken. Wahrscheinlich ist der Unterschied im Quarzitgrus-Gehalt dafür verantwortlich. Wegen der relativ grusreichen Oberschicht des Substrats trocknen die Böden der Variante mit Galium vivianum wahrscheinlich weniger schnell aus als die relativ sandreicheren Böden der Variante mit Corynephorus canescens. In allen Ausbildungen wird die Beweglichkeit des Materials ohne Zweifel durch Abspülung (Regenwasser, Schmelzwasser), Winderosion (Ablation/Deflation) und Kammeisbildung verursacht, deren Einflüße an den Hängen von der Schwerkraft verstärkt werden. Wie diese Prozesse genau verlaufen, ist unbekannt; die Erforschung der dynamischen hydro-äolischen und kryologischen Prozesse innerhalb bestimmter Vegetationstypen (z.B. im Violetum langeanae) wird aber zur Zeit untersucht..
Die an primären und sekundären Standorten herrschenden ökologischen Bedingungen (Beweglichkeit des Materials) stimmen einigermaßen mit denen der reinen Schuttfluren überein. Die Gesellschaft ist am besten im Spätfrühling entwickelt. Die gelben Blüten von Viola langeana sind in den niederen Lagen des Gebirges schon ab März zu beobachten. Die Art dürfte nach Beobachtungen von Jan JANSEN in der Estrela bis auf 800 m NN (vor allem an Wegböschungen) hinabreichen. Die Gesellschaft entwickelt sich sowohl in der supra- als auch in der oromediterranen Zone, scheint aber am besten zwischen 1600 und 1800 m Höhe zu gedeihen.
 | |
 |  |
 |  |
 |  |
 |  |
 |  |
 |  |
3. Höhlen
Höhlen und ehemalige Bergwerke sind sehr wichtige Lebensräume für Fledermäuse, von denen bis zu 19 Arten innerhalb der Grenzen des Parks auftreten.
Diese Umgebungen sind auch von botanischem Interesse, da sie Farne und Moose beherbergen. Die Farnarten sind von anderen Vegetationstypen her bekannt und werden hier nicht berücksichtigt. Ein auffälliger Bryophyt wird hier jedoch angesprochen, nämlich das Leuchtmoos Schistostega pennata. Dieses Moos erscheint lumineszent, verursacht durch die lichtreflektierenden Eigenschaften des Protonemas, einem Vorkeim im Lebenszyklus von Bryophyten. Schistostega pennata bildet bläulich-grün leuchtende Flecken an dunklen Orten wie Eingänge zu Höhlen, alten Bergwerken und Felsen, die von Hirten für eine Benutzung als Unterstände ausgegraben wurden.


{kind=link}
{kind=link}
Natura 2000 Geröllhalden-Habitate
Der Habitat 8130 der Geröllhalden (Schutthalden) großer Höhen bei oro-temperatem Bioklima ist für Portugal (Serra da Estrela) im ‘Plano Sectorial’ von Natura 2000 (ICNF) beschrieben:
- Granitische Geröllhalden bei supratempertem bis orotemperatem Bioklima mit Verbreitung in der Serra da Estrela.
- Die häufigsten Gefässpflanzen in den Geröllhalden sind mehrjährige Pflanzen, häufig mit Stolonen, rhizomatös oder mit einem ausgedehnten Wurzelsystem, das sich in die Tiefe parallel zur Oberfläche erstreckt. Sie werden phytosoziologisch als charakteristische Arten der Klasse Thlaspietea rotundifolii angesehen, u.a.: Arrhenatherum elatius subsp. carpetanus, Coincya monensis subsp. orophila, Digitalis purpurea subsp. carpetana, Doronicum carpetanum, Dryopteris expansa, D. oreades, Eryngium duriaei subsp. duriaei, Lactuca viminea subsp. viminea, Leontodon hispidus subsp. bourgaeanus, Linaria saxatilis subsp. saxatilis, Paronychia polygonifolia, Phalacrocarpum oppositifolium subsp. oppositifolium, Reseda gredensis, Rumex suffruticosus, Scrophularia herminii, Senecio pyrenaicus subsp. caespitosus, Silene foetida subsp. foetida, Solidago virgaurea subsp. fallit-tirones, Trisetaria hispida.
- In den Gerollhalden finden sich häufig noch Elemente der Borstgrasrasen (siehe Habitat 6230 ‘Artenreiche Nardus-Formationen auf silikatreichen Böden in alpinen Bergzonen (und subalpinen Höhenstufen in Zentraleuropa)’) sowie die psicroxerophilen Rasen (Habitat 6160 der ‘Oro-iberischen Festuca indigesta Rasen’).
- In den orphilen silikatreichen Schutthalden wurden drei Phytozönosen der Thlaspietea rotundifolii mit unterschiedlichen Ansprüchen, was die Wasserversorgung und die Mobilität und Dimension der Schuttfragmente anbetrifft, bestimmt.
- In der Serra da Estrela kann die Vegetation der Thlaspietea rotundifolii stellenweise auch in Moränen und Blockhalden auftreten, was ihre Interpretation im Bereich dieses Subtypus erforderlich macht.
Phytosoziologische Korrespondenz
- Die Ordnungen Androsacetalia alpinae e Polystichetalia lonchitidis innerhalb der Klasse Thlaspietea rotundifolii.
Bioindikatoren
- Präsenz in variablen floristischen Kompositionen von Cryptogramma crispa, Digitalis purpurea subsp. carpetana, Dryopteris oreades, Eryngium duriaei subsp. duriaei, Leontodon hispidus subsp. bourgaeanus, Phalacrocarpum oppositifolium subsp. oppositifolium, Reseda gredensis, Rumex suffruticosus, Scrophularia herminii, Senecio pyrenaicus subsp. caespitosus, Silene foetida subsp. foetida, Trisetaria hispida.
Annexe
1 Phytosoziologische Aufnahmen
Synoptische Tabellen der Vegetationsaufnahmen der Klasse Thlaspietea zur Schutthaldenvegetation in der Serra da Estrela. Entnommen aus: Jan JANSEN (2011): Managing Natura 2000 in a changing world: - Radboud Universiteit - Appendix 1 Vegetation tables (Chapter 5, Table 1 and Table 2). (Hier gelb unterlegt: Flechten und Moose).
Annex 1 - Tabelle 1
Chapter 5, Table 1: Übersicht der Thlaspietea-Gesellschaften in den höheren Stufen der Serra da Estrela
     |
 |
Annex 1 - Tabelle 2
Chapter 5, Table 2 Violetum langeanae Ortiz & Samaniego 1989
   |
2 Phytosoziologische Syntaxa
2.1 Syntaxa zu der Geröllhaldenvegetation THLASPIETEA ROTUNDIFOLII
(Aus: Jan JANSEN (2002): Geobotanical guide of the Serra da Estrela. Instituto da Conservação da Natureza. Ministério das Cidades, Ordenamento do Território e Ambiente, Lisboa.und Jan JANSEN (2011): Managing Natura 2000 in a changing world: - Radboud Universiteit .
Die Beschreibungen zu den Phytosoziologischen Syntaxa sind RIVAS-MARTÍNEZ et. al. (2002): Vascular Plant Communities of Spain and Portugal. Addenda to the Syntaxonomical Checklist of 2001. ITINERA GEOBOTANICA. Vol. 15(1). entnommen)

(Aus: Jan Jansen (2002) :Geobotanical guide of the Serra da Estrela. Instituto da Conservação da Natureza. Ministério das Cidades, Ordenamento do Território e Ambiente, Lisboa.)

(Aus: Chapter 5 in: Jansen (2011): Managing Natura 2000 in a changing world: - Radboud Universiteit .)
Klasse THLASPIETEA ROTUNDIFOLII Br.-Bl. 1948
Loose shifting scree, stone and pebble perennial communities of mountain slopes, moraines and riverine gravel deposits, growing on boreal, temperate and mediterranean bioclimates in the Holarctic territories.
Temperate and Mediterranean high calcareous mountains, calc-schist or calcium carbonate rich shifting stones (Thlaspietalia rotundifolii) [oro-cryorotemperate Pyrenean (Iberidion spathulatae), oro-cryorotemperate Orocantabric (Linarion filicaulis), Pyrenean and Orocantabric wet and long snow covered (Saxifragion praetermissae), supratemperate
Orocantabric and Castilian Cantabrian (Iberido-Linarion propinquae), supraoromediterranean
Betic (Platycapno-Iberidion lagascanae)]; Temperate and Mediterranean high siliceous mountains and lacking calcium carbonate shifting stones (Androsacetalia alpinae) [oro-cryorotemperate Pyrenean (Senecionion leucophylli), meso-supratemperate Pyrenean (Galeopsion pyrenaicae), Carpetan, Bercian-Sanabriensean, Oroiberian and Orocantabric (Linario saxatilis-Senecionion carpetani), Nevadensian oro-cryoromediterranean (Holcion caespitosi)]; boreal, temperate and mediterranean high mountain deposits of blocks by periglacial congelifraction, glacial erratic, etc., rich in ferns (Polystichetalia lonchitidis) [rich in calcium carbonate (Gymnocarpion robertiani), lacking calcium carbonate (Dryopteridion oreadis)]; thermo to supramediterranean dry and subhumid gravel, pebble and block slopes in Western Mediterranean (Andryaletalia ragusinae) [riverine deposits (Glaucion flavi), Setabensean and Valencian shifting stones and blocks (Scrophularion sciophilae)]; supra and lower orotemperate and submediterranean Alpine, Middle European, Pyrenean, Orocantabric and Oroiberian calcareous or calcium carbonate rich loose scree and riverine gravel deposits (Achnatheretalia calamagrostis) [loose shifting stones (Achnatherion calamagrostis), Pyrenean, Orocantabric and Oroiberian riverine (Calamagrostion pseudophragmitis)]; upper cryorotemperate early snow-free on poor, neutral or rich screes or congelifracted slopes of Alpine and Pyrenean high mountains (Drabetalia hoppeanae) [Pyrenean nival summits (Androsacion ciliatae)]. European Holarctic.
(Aus: Rivas-Martínez et. al. (2002): Vascular Plant Communities of Spain and Portugal. Addenda to the Syntaxonomical Checklist of 2001. ITINERA GEOBOTANICA. Vol. 15(1).)
|
Jan JANSEN (2011) verzichtet wegen des noch unzureichneneden Kenntnisstandes auf eine phytosoziologische Zuordnung des Violetum langianae. COSTA et al. 2012 ordnet das Violetum langianae der Klasse Thlaspietea zu. Weitere Untersuchungen zu dieser Gesellschaft zur weiteren Klärung ihrer phytosoziologischen Stellung sind im Gange.
| |
| |
2.2 Syntaxa zu den übrigen Felsbiotopen der Serra da Estrela (Jan JANSEN, 2002)
|
8230 Silikatfelsen mit Pioniervegetation des Sedo-Scleranthion oder des Sedo albi-Veronicion dillenii
|
 |
 |
 |
  |
 |
 |
(Fussnoten und Bibliographie: Da sich Fussnoten einschl. Bibliographie nicht ohne weiteres aus Google-Docs heraus nach Blogger übertragen lassen, können diese im veröffentlichten Originalartikel eingesehen werden -> https://docs.google.com/document/d/1NrHbQigYKM47lu0ztN0Hxd_n403Z-Q0o31aQoy21f9A/pub

No comments:
Post a Comment